

Soil AMF community structure and assembly mechanism of medical sativa field in Loess Plateau
-
摘要: 为揭示多年种植苜蓿对土壤丛枝菌根真菌(AMF)群落结构和多样性的影响, 本研究通过布设在黄土高原半干旱区的田间试验, 基于2019、2012和2003年建植的紫花苜蓿(Medicago sativa), 以农田玉米(Zea mays)为对照, 采用高通量测序和PCR技术, 结合分子生态网络研究不同种植年限紫花苜蓿地土壤AMF群落组成和丰度, 并基于零模型揭示了土壤AMF群落的组装过程。结果表明: 黄绵土区AMF属于球囊菌门的1纲4目7科7属, 球囊霉属、类球囊霉属和多孢囊霉属为紫花苜蓿地和农田土壤共有类群, 且均以球囊霉属(65.15%~99.12%)为优势属, 其主要贡献了不同处理分组中土壤AMF群落结构的改变。长期种植紫花苜蓿使得和平囊霉属和无梗囊霉属消亡, 但促生了双型囊霉属和盾巨孢囊霉属, 其中双型囊霉属相对丰度表现为L2019处理显著高于其他处理(P<0.05)。网络关联分析发现, 高丰度的球囊霉属和类球囊霉属之间呈现负相关, 而低丰度的和平囊霉属和无梗囊霉之间呈现正相关。基于零模型的群落组装结果表明, 农田与L2019处理由确定性过程主导(66.67%), L2012和L2003处理由随机性过程主导(100%), 这表明长期种植紫花苜蓿形成稳定的土壤环境使其随机性过程增加, 利于维持人工草地生态系统功能的可持续性和稳定性。Abstract: Arbuscular mycorrhizal fungi (AMF) mediate interactions between plants and soils, which play crucial role in terrestrial symbiosis and one of the important components in soil microbial community. However, little is known about how soil AMF community varies in relation to soil properties loessial soil. Therefore, the present study investigated soil AMF diversity community structure and soil physicochemical properties in Medicago sativa field and Farmland in the semi-arid area of the Loess Plateau. Soil samples (0-20 cm) were taken from four treatments in June 2021:1) farmland (Zea mays) and Medicago sativa established in 2019(L2019), 2012(L2012), 2003(L2003). The Illumina MiSeq high-throughput sequencing and real-time fluorescent quantitative PCR technology were used to explore the structure and diversity of AMF communities under four treatments (Farmland, L2003, L2012 and L2019). The statistical methods (such as redundancy analysis and molecular ecological network analysis) were used to explore the relationship between soil physicochemical properties and AMF community. Zero model analysis were used to reveal the assembly process of soil AMF community. The results showed that long-term planting of alfalfa decreased soil total phosphorus and available phosphorus. The AMF gene abundance ranged from 1.02×104~copies∙g−1 to 1.50×104copies·g-1 in dry soil, being significantly higher in Medicago sativa established in 2003(L2003) than other treatment (P<0.05). The correlation analysis between the abundance of AMF gene and physicochemical factors showed that soil AMF gene abundance was positively correlated with Total nitrogen, and negatively correlated with total phosphorus content, and available phosphorus. A total of 1 class, 4 orders, 7 families, and 7 genera of AMF were identified. Glomus, Diversispora and Paraglomus were the common genera of Medicago sativa field and Farmland, the dominant genera of Medicago sativa field and Farmland were Glomus (65.15%−99.12%), It mainly contributed to the changes of soil AMF community structure in different treatment groups. Long-term cultivation of Medicago sativa propagated the rare microbial taxa, including Ambispora and Scutellospora, whereas Pacispora and Acaulospora were sterilized. Ambispora Were significantly higher in Medicago sativa established in 2019(L2019) than other treatment (P<0.05), The analysis of molecular ecological network showed that there were high-abundance genera (Glomus and Paraglomus) had a cooperative relationship in the ecological network, while the low-abundance genera (Pacispora and Acaulospora )had a competitive relationship in the ecological network. RDA analysis showed that there was no main environmental factor affecting the community structure of AMF. Null model was used to infer AMF community assembly processes. Farmland and Medicago sativa established in 2019 (L2019) communities mechanism were dominantly assembled with the deterministic process (66.67%), the heterogeneous selection contributed the most. Medicago sativa established in 2012 (L2012) and 2003 (L2003) communities mechanism were dominantly assembled with random process (100%), the undominated processes contributed the most in Medicago sativa established in 2012 (L2012) and the dispersal limitation contributed the most in Medicago sativa established in 2003 (L2003). Mantel test showed that there was no main driving environmental factor for AMF community assembly. Long-term Medicago sativa cultivation increased the random processes. It is beneficial to maintain the sustainability and stability of artificial grassland ecosystem function. In summary, long-term planting of Medicago sativa significantly affected the composition of AMF community in soil. This study provides basic data and theoretical basis for further study on the microbial mechanism of AMF in the Loess Plateau after years of Medicago sativa planting.
-
Key words:
- Medicago sativa /
- Arbuscular mycorrhizal fungi /
- Community structure /
- Community assembly /
- Loessal soil
-

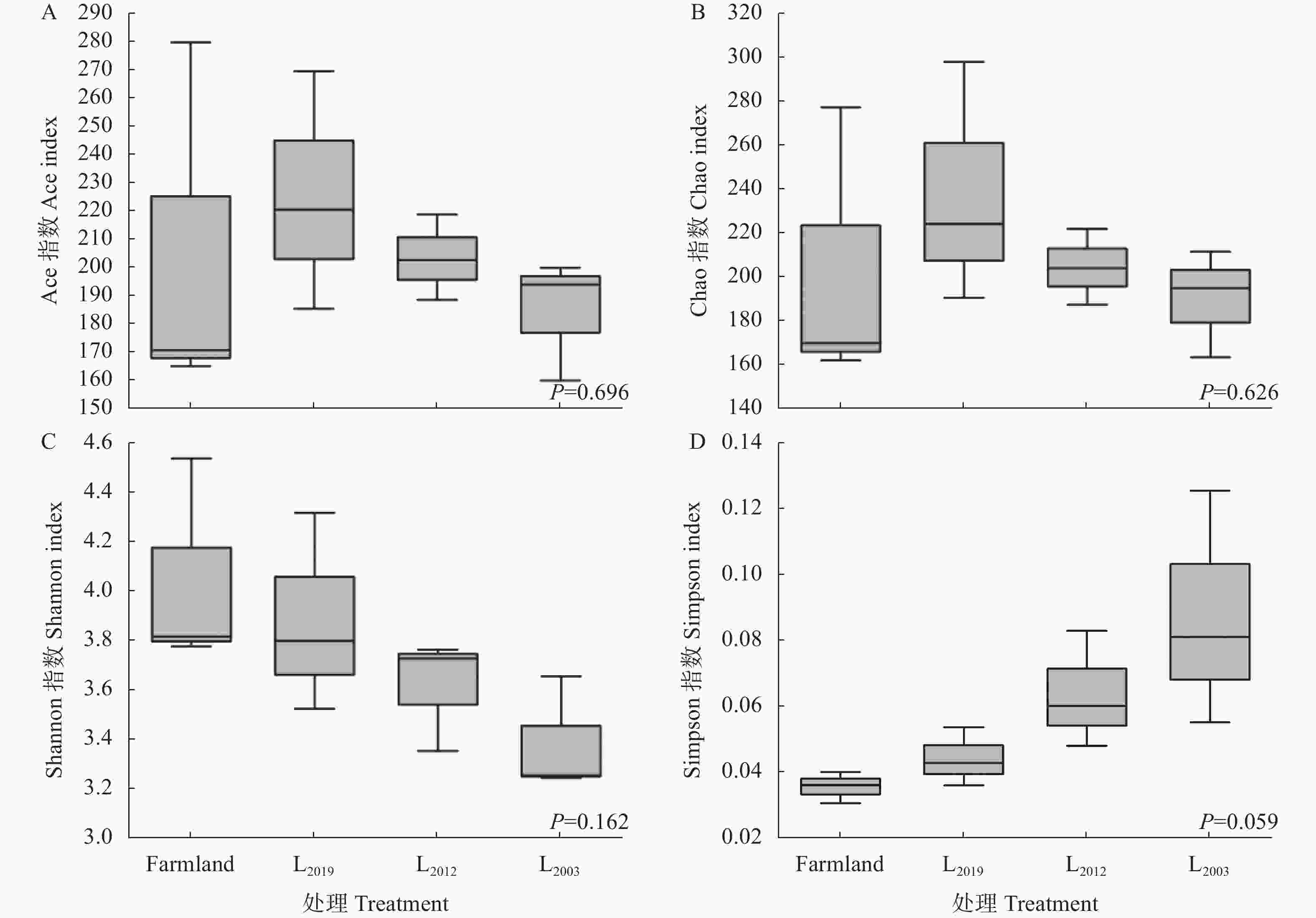
图 1 不同处理土壤AMF群落多样性指数
Farmland、L2019、L2012、L2003分别表示农田、2019年建植苜蓿、2012 年建植苜蓿和2003年建植苜蓿。Farmland, L2019, L2012, and L2003 denote farmland and Medicago sativa planting years of 2019, 2012, and 2003, respectively.
Figure 1. Diversity indexs of soil AMF community under different treatments
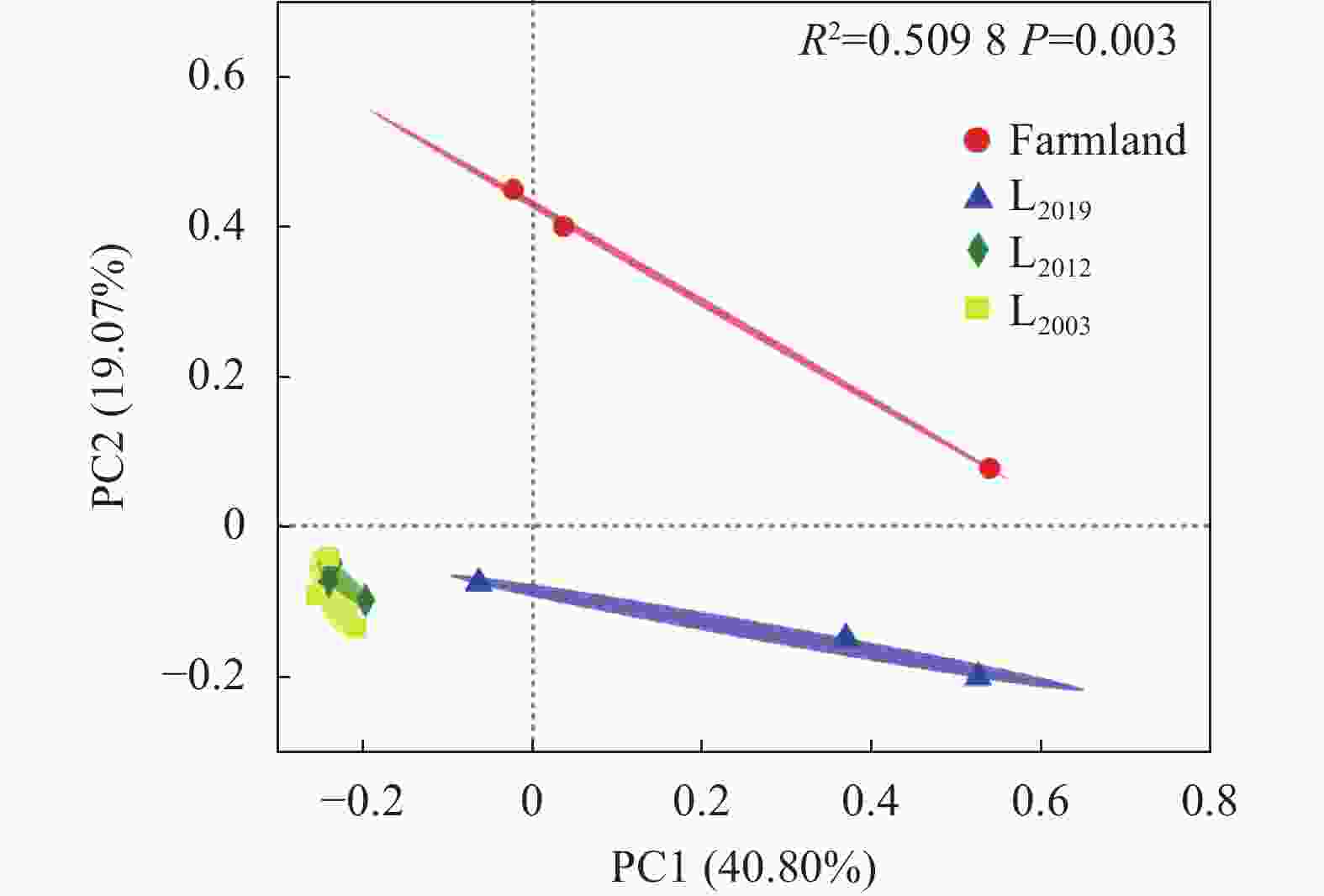
图 2 不同处理土壤AMF群落主坐标分析
Farmland、L2019、L2012、L2003分别表示农田、2019年建植苜蓿、2012 年建植苜蓿和2003年建植苜蓿。Farmland, L2019, L2012, and L2003 denote farmland and Medicago sativa planting years of 2019, 2012, and 2003, respectively.
Figure 2. Principal co-ordinates analysis (PcoA) of soil AMF communities under different treatments

图 3 不同处理土壤AMF属水平群落结构(A)和双型囊霉属相对丰度(B)
Farmland、L2019、L2012、L2003分别表示农田、2019年建植苜蓿、2012 年建植苜蓿和2003年建植苜蓿。Farmland, L2019, L2012, and L2003 denote farmland and Medicago sativa planting years of 2019, 2012, and 2003, respectively.
Figure 3. Soil AMF community at the genus levels (A) and relative abundance of Ambispora (B) under different treatments
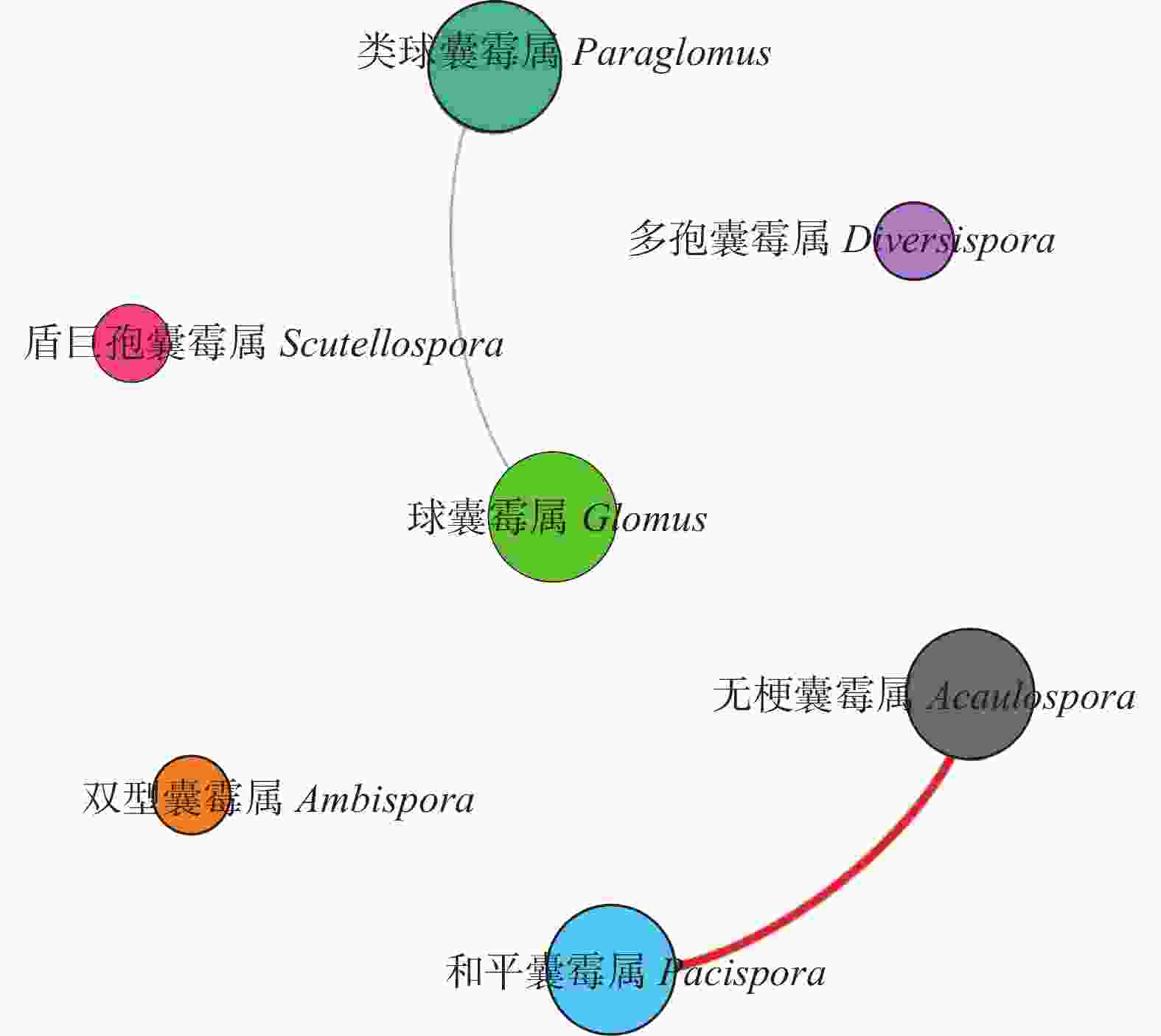
图 4 农田和紫花苜蓿地AMF属的关联网络图
红线表明正相关, 绿线表明负相关。Red lines represent positive correlation. Green lines represent negative correlation.
Figure 4. Associated network at different AMF genus under Farmland and Medicago sativa field
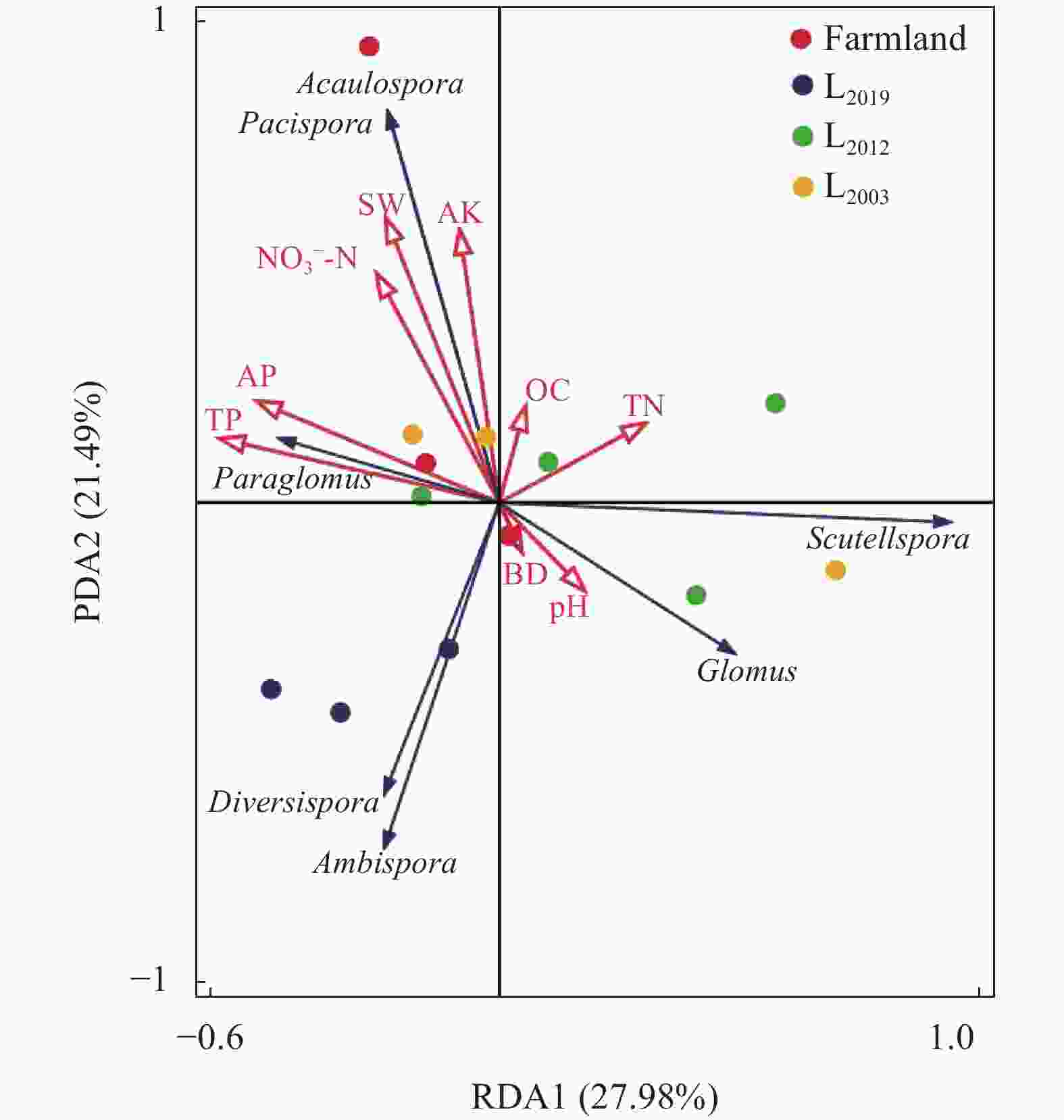
图 5 土壤AMF群落与土壤理化因子冗余分析
Farmland、L2019、L2012、L2003分别表示农田、2019年建植苜蓿、2012 年建植苜蓿和2003年建植苜蓿。Glomus, Paraglomus, Diversispora, Scutellospora, Ambispora, Acaulospora and pacispora分别表示球囊霉属、类球囊霉属、多孢囊霉属、盾巨孢囊属、双型囊霉属、无梗囊霉属和平囊霉属。Farmland, L2019, L2012, and L2003 denote farmland and Medicago sativa planting years of 2019, 2012, and 2003, respectively.
Figure 5. Redundancy analysis (RDA) of soil AMF communities and soil physicochemical properties
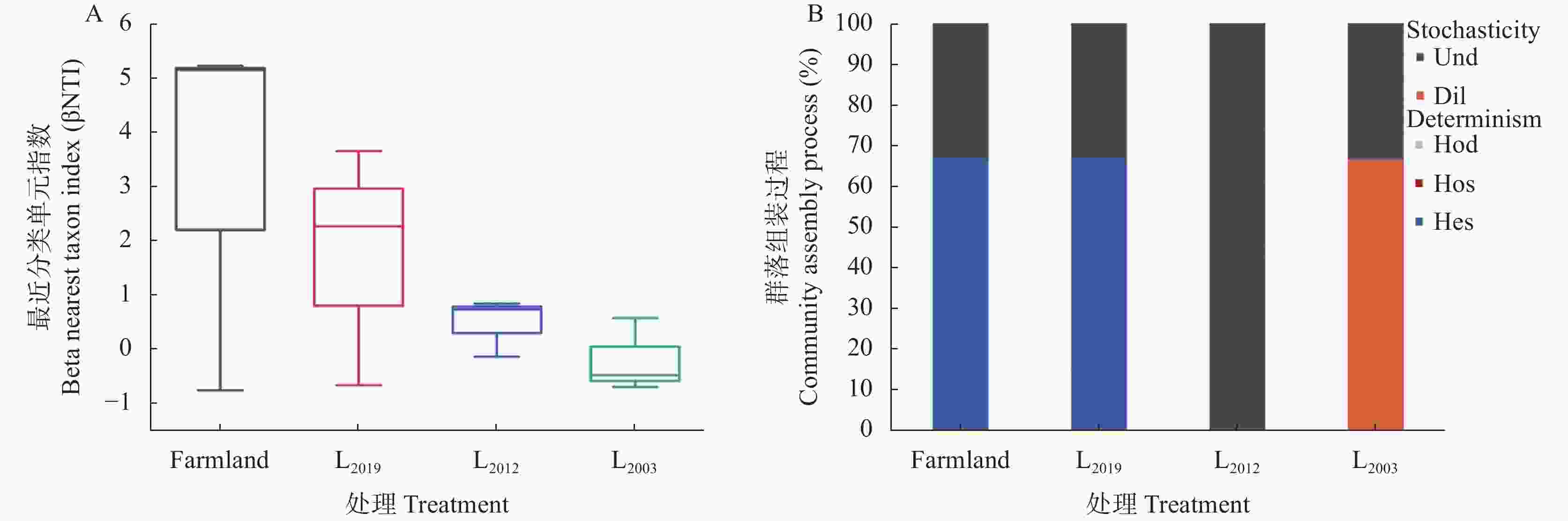
图 6 不同处理土壤AMF群落组装生态过程
Farmland、L2019、L2012、L2003分别表示农田、2019年建植苜蓿、2012 年建植苜蓿和2003年建植苜蓿。Stochasticity: 随机性过程; Dil: 扩散限制; Hod: 同质性扩散; Und: 非主导过程; Determinism: 确定性过程; Hes: 异质性选择; Hos: 同质性选择。Farmland, L2019, L2012, and L2003 denote farmland and Medicago sativa planting years of 2019, 2012, and 2003, respectively. Dil: dispersal limitation; Hod: homogenizing dispersal; Und: undominated processes; Hes: heterogeneous selection; Hos: homogeneous selection.
Figure 6. Ecological processes governing soil AMF community assembly under different treatment
表 1 不同处理土壤基本理化性状及AMF基因丰度
Table 1. Soil physicochemical properties and Abundance of AMF gene under different treatments
指标 Index Farmland L2019 L2012 L2003 土壤水分 Soil water (%) 15.65±0.76a 8.76±0.46b 8.72±0.34b 9.62±0.13b 容重 Bulk Density (g∙cm−3) 1.18±0.01a 1.21±0.01a 1.23±0.02a 1.24±0.03a 有机碳 Organic carbon (g∙kg−1) 10.50±0.20b 9.83±0.21b 9.89±0.05b 11.49±0.32a 全氮 Total nitrogen (g∙kg−1) 0.92±0.03b 0.77±0.02c 0.85±0.05bc 1.11±0.05a 硝态氮 Nitrate nitrogen (mg∙kg−1) 23.14±0.33a 13.93±0.07c 12.35±0.07d 14.85±0.08b 全磷 Total phosphorus (g∙kg−1) 0.99±0.01a 0.93±0.01b 0.86±0.03c 0.82±0.01c 速效磷 Available phosphorus (mg∙kg−1) 6.21±0.05a 5.02±0.15b 3.92±0.13c 3.54±0.09d 速效钾 Available potassium (mg∙kg−1) 223.00±11.14a 222.00±2.89a 228.67±7.06a 229.33±0.88a pH 8.38±0.03a 8.49±0.01a 8.47±0.04a 8.45±0.03a AMF基因丰度
Abundance of AMF gene [×104 copy∙g−1(dry soil)]1.15±0.01bc 1.02±0.00c 1.26±0.04b 1.50±0.08a 数据为平均值±标准误(n=3), 同行不同小写字母表示不同处理间差异显著(P<0.05), Farmland、L2019、L2012、L2003分别表示农田、2019年建植苜蓿、2012年建植苜蓿和2003年建植苜蓿。Data in table are mean ± standard error (n=3). Different lowercase letters in the same line indicate significant difference between different treatments (P<0.05). Farmland, L2019, L2012, and L2003 denote farmland and Medicago sativa planting years of 2019, 2012, and 2003, respectively.  下载: 导出CSV
下载: 导出CSV
表 2 AMF基因丰度与土壤理化因子相关分析
Table 2. Correlation analysis of AMF gene abundance and environmental factors
指标 Index Abundance of AMF gene SW BD OC TN NO3−-N TP AP AK pH Abundance of AMF gene 1 SW 0.105 1 BD 0.298 −0.495 1 OC 0.522 0.522 0.165 1 TN 0.789** 0.474 0.023 0.786** 1 NO3--N 0.007 0.888** −0.488 0.648* 0.523 1 TP −0.711** 0.438 −0.627* −0.202 −0.304 0.529 1 AP −0.774** 0.326 −0.560 −0.263 −0.397 0.396 0.961** 1 AK 0.462 −0.042 0.229 0.109 0.305 −0.112 −0.385 −0.392 1 pH −0.102 −0.615* 0.456 −0.252 −0.515 −0.566 −0.327 −0.322 0.162 1 **: P<0.01; *: P<0.05. 表中Abundance of AMF gene、SW、BD、OC、TN、NO3−-N、TP、AP、AK、pH 分别表示AMF基因丰度、土壤水分、容重、有机碳、全氮、硝态氮、全磷、速效磷、速效钾、pH。In the table, Abundance of AMF gene, SW, BD, SOC, TN, NO3−-N, TP, AP, AK, pH were soil water, bulk Density, organic carbon, total nitrogen, nitrate nitrogen, total phosphorus, available phosphorus, available potassium, pH.
下载: 导出CSV
表 3 不同处理方式间AMF群落组成差异的优势属贡献率
Table 3. Contributions of dominant species to AMF community compositions under different treatment
% 分组
Groups球囊霉属
Glomus类球囊霉属
Paraglomus多孢囊霉属
Diversispora盾巨孢囊属
Scutellospora双型囊霉属
Ambispora无梗囊霉属
Acaulospora和平囊霉属
pacisporaFarmland vs L2019 48.08 5.27 1.23 0.00 0.93 0.18 0.18 Farmland vs L2012 49.00 1.91 0.56 1.39 0.35 0.27 0.27 Farmland vs L2003 49.18 2.64 0.64 1.14 0.06 0.27 0.26 L2019 vs L2012 49.16 6.06 1.53 0.99 0.74 0.00 0.00 L2019 vs L2003 49.26 5.64 1.32 0.84 0.96 0.00 0.00 L2012 vs L2003 40.57 27.44 5.50 21.19 4.45 0.00 0.00 Farmland、L2019、L2012、L2003分别表示农田、2019年建植苜蓿、2012 年建植苜蓿和2003年建植苜蓿。Farmland, L2019, L2012, and L2003 denote farmland and Medicago sativa planting years of 2019, 2012, and 2003, respectively.
下载: 导出CSV
表 4 土壤理化因子与βNTI的Mantel分析
Table 4. Mantel tests of soil physicochemical properties and βNTI
因子 Factor βNTI r P SW −0.126 0.818 BD −0.131 0.846 OC 0.109 0.198 TN −0.102 0.754 NO3−-N −0.071 0.651 TP −0.056 0.627 AP −0.124 0.843 AK −0.032 0.518 pH 0.099 0.242
下载: 导出CSV
-
[1] PAN S, WANG Y, QIU Y P, et al. Nitrogen-induced acidification, not N-nutrient, dominates suppressive N effects on arbuscular mycorrhizal fungi[J]. Global Change Biology, 2020, 26(11): 6568−6580 doi: 10.1111/gcb.15311 [2] LUKAS S, GATTINGER A, MEIER M, et al. Improving Crop Yield and Nutrient Use Efficiency via Biofertilization-A Global Meta-analysis[J]. Frontiers in Plant Science, 2018, 8(12): 2204 [3] VOLPIN H, PHILLIPS D A. Respiratory elicitors from Rhizobium meliloti affect intact alfalfa roots[J]. Plant Physiology, 1998, 116(2): 777−783 doi: 10.1104/pp.116.2.777 [4] MASSA N, CESARO P, TODESCHINI V, et al. Selected autochthonous rhizobia, applied in combination with AM fungi, improve seed quality of common bean cultivated in reduced fertilization condition[J]. Applied Soil Ecology, 2020, 148: 103507 doi: 10.1016/j.apsoil.2020.103507 [5] 赵丹丹, 李涛, 赵之伟. 丛枝菌根真菌-豆科植物-根瘤菌共生体系的研究进展[J]. 生态学杂志, 2006, 25(3): 327−333 doi: 10.3321/j.issn:1000-4890.2006.03.020ZHAO D D, LI T, ZHAO Z W. Research advances in arbuscular mycorrhizal fungi-legumes-rhizobia symbiosis[J]. Chinese Journal of Ecology, 2006, 25(3): 327−333 doi: 10.3321/j.issn:1000-4890.2006.03.020 [6] ABD-ALLA M H, ELSADEK EL-ENANY A W, ALLAM NAFADY N, et al. Synergistic interaction of Rhizobium leguminosarum bv. viciae and arbuscular mycorrhizal fungi as a plant growth promoting biofertilizers for faba bean (Vicia faba L. ) in alkaline soil[J]. Microbiological Research, 2014, 169(1): 49−58 doi: 10.1016/j.micres.2013.07.007 [7] 李元敬, 刘智蕾, 何兴元, 等. 丛枝菌根共生体的氮代谢运输及其生态作用[J]. 应用生态学报, 2013, 24(3): 861−868LI Y J, LIU Z L, HE X Y, et al. Nitrogen metabolism and translocation in arbuscular mycorrhizal symbiote and its ecological implications[J]. Chinese Journal of Applied Ecology, 2013, 24(3): 861−868 [8] CHALK P M, DE F SOUZA R, URQUIAGA S, et al. The role of arbuscular mycorrhiza in legume symbiotic performance[J]. Soil Biology and Biochemistry, 2006, 38(9): 2944−2951 doi: 10.1016/j.soilbio.2006.05.005 [9] 伏云珍, 马琨, 崔慧珍, 等. 间作作物种间相互作用对马铃薯根际土壤丛枝菌根真菌的影响[J]. 生态学杂志, 2021, 40(1): 131−139 doi: 10.13292/j.1000-4890.202101.030FU Y Z, MA K, CUI H Z, et al. Effects of interspecific interactions between intercropping crops on arbuscular mycorrhizal fungi in potato rhizosphere soil in the intercropping system[J]. Chinese Journal of Ecology, 2021, 40(1): 131−139 doi: 10.13292/j.1000-4890.202101.030 [10] LEIBOLD M A, MCPEEK M A. Coexistence of the niche and neutral perspectives in community ecology[J]. Ecology, 2006, 87(6): 1399−1410 doi: 10.1890/0012-9658(2006)87[1399:COTNAN]2.0.CO;2 [11] 柴宇星. 祁连山阴阳坡丛枝菌根真菌多样性及群落构建机制研究[D]. 兰州: 兰州大学, 2018CHAI Y X. The species diversity and community assembly of arbuscular mycorrhizal fungi in Qilianshan Mountains on A northwest-facing slope and A southeast-facing slope[D]. Lanzhou: Lanzhou University, 2018 [12] VÁLYI K, MARDHIAH U, RILLIG M C, et al. Community assembly and coexistence in communities of arbuscular mycorrhizal fungi[J]. The ISME Journal, 2016, 10(10): 2341−2351 doi: 10.1038/ismej.2016.46 [13] 李洁, 李杏春, 郭良栋. 真菌群落构建机制研究进展[J/OL]. 菌物学报: 1–21 [2023–01–26]. http://kns.cnki.net/kcms/detail/11.5180.Q.20221122.1836.008.htmlLi J, Li X C, Guo L D. Research progress on community assembly mechanisms of fungi[J/OL]. Mycosystema: 1–21 [2023–01–26]. http://kns.cnki.net/kcms/detail/11.5180.Q.20221122.1836.008.html [14] 倪红, 杨宪龙, 王刚, 等. 施氮及添加硝化抑制剂对苜蓿草地N2O排放的影响[J]. 中国生态农业学报(中英文), 2020, 28(3): 317−327NI H, YANG X L, WANG G, et al. Effects of nitrogen application and nitrification inhibitor addition on N2O emissions in Medicago sativa L. grassland[J]. Chinese Journal of Eco-Agriculture, 2020, 28(3): 317−327 [15] 江影舟, 南志标, 王丽佳. 基于钻石模型理论的甘肃省苜蓿产业竞争力分析[J]. 草业科学, 2016, 33(4): 813−820 doi: 10.11829/j.issn.1001-0629.2015-0363JIANG Y Z, NAN Z B, WANG L J. Competitiveness of alfalfa industry in Gansu Province—a diamond model[J]. Pratacultural Science, 2016, 33(4): 813−820 doi: 10.11829/j.issn.1001-0629.2015-0363 [16] WANG L L, XIE J H, LUO Z Z, et al. Forage yield, water use efficiency, and soil fertility response to alfalfa growing age in the semiarid Loess Plateau of China[J]. Agricultural Water Management, 2021, 243: 106415 doi: 10.1016/j.agwat.2020.106415 [17] Recorbet G, Calabrese S, Balliau T, et al. Proteome adaptations under contrasting soil phosphate regimes of Rhizophagus irregularis engaged in a common mycorrhizal network[J]. Fungal Genetics and Biology, 2021, 147: 103517 doi: 10.1016/j.fgb.2021.103517 [18] ZHANG L, FENG G, DECLERCK S. Signal beyond nutrient, fructose, exuded by an arbuscular mycorrhizal fungus triggers phytate mineralization by a phosphate solubilizing bacterium[J]. The ISME Journal, 2018, 12(10): 2339−2351 doi: 10.1038/s41396-018-0171-4 [19] ZHANG L, FAN J Q, DING X D, et al. Hyphosphere interactions between an arbuscular mycorrhizal fungus and a phosphate solubilizing bacterium promote phytate mineralization in soil[J]. Soil Biology and Biochemistry, 2014, 74: 177−183 doi: 10.1016/j.soilbio.2014.03.004 [20] 杨文莹, 孙露莹, 宋凤斌, 等. 陆地农业生态系统丛枝菌根真菌物种多样性研究进展[J]. 应用生态学报, 2019, 30(11): 3971−3979YANG W Y, SUN L Y, SONG F B, et al. Research advances in species diversity of arbuscular mycorrhizal fungi in terrestrial agro-ecosystem[J]. Chinese Journal of Applied Ecology, 2019, 30(11): 3971−3979 [21] 张旭红, 朱永官, 王幼珊, 等. 不同施肥处理对丛枝菌根真菌生态分布的影响[J]. 生态学报, 2006, 26(9): 3081−3087 doi: 10.3321/j.issn:1000-0933.2006.09.038ZHANG X H, ZHU Y G, WANG Y S, et al. Effect of long-term fertilization on the diversity and distribution of arbuscular mycorrhiza fungi in Northeast China[J]. Acta Ecologica Sinica, 2006, 26(9): 3081−3087 doi: 10.3321/j.issn:1000-0933.2006.09.038 [22] WILLIAMS A, MANOHARAN L, ROSENSTOCK N P, et al. Long-term agricultural fertilization alters arbuscular mycorrhizal fungal community composition and barley (Hordeum vulgare) mycorrhizal carbon and phosphorus exchange[J]. The New Phytologist, 2017, 213(2): 874−885 doi: 10.1111/nph.14196 [23] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2000BAO S D. Soil and Agricultural Chemistry Analysis[M]. 3rd ed. Beijing: China Agriculture Press, 2000. [24] STEGEN J C, LIN X J, FREDRICKSON J K, et al. Quantifying community assembly processes and identifying features that impose them[J]. The ISME Journal, 2013, 7(11): 2069−2079 doi: 10.1038/ismej.2013.93 [25] 刘润进, 李晓林. 丛枝菌根及其应用[M]. 北京: 科学出版社, 2000LIU R J, LI X L. Application of arbuscular mycorrhizal [M]. Beijing: Science Press, 2000 [26] WU M N, QIN H L, CHEN Z, et al. Effect of long-term fertilization on bacterial composition in rice paddy soil[J]. Biology and Fertility of Soils, 2011, 47(4): 397−405 doi: 10.1007/s00374-010-0535-z [27] URCOVICHE R C, GAZIM Z C, DRAGUNSKI D C, et al. Plant growth and essential oil content of Mentha crispa inoculated with arbuscular mycorrhizal fungi under different levels of phosphorus[J]. Industrial Crops and Products, 2015, 67: 103−107 doi: 10.1016/j.indcrop.2015.01.016 [28] 吴强盛, 夏仁学, 邹英宁. 柑橘丛枝菌根真菌生长与根际有效磷和磷酸酶活性的相关性[J]. 应用生态学报, 2006, 17(4): 4685−4689 doi: 10.3321/j.issn:1001-9332.2006.04.025WU Q S, XIA R X, ZOU Y N. Arbuscular mycorrhizal fungal growth on citrus roots and its correlations with soil available phosphorus content and phosphatase activity[J]. Chinese Journal of Applied Ecology, 2006, 17(4): 4685−4689 doi: 10.3321/j.issn:1001-9332.2006.04.025 [29] 王永明, 范洁群, 石兆勇. 中国丛枝菌根真菌分子多样性[J]. 微生物学通报, 2018, 45(11): 2399−2408 doi: 10.13344/j.microbiol.china.171093WANG Y M, FAN J Q, SHI Z Y. Molecular diversity of arbuscular mycorrhizal fungal in China[J]. Microbiology China, 2018, 45(11): 2399−2408 doi: 10.13344/j.microbiol.china.171093 [30] 薛壮壮, 冯童禹, 王超, 等. 土地利用方式对酸性红壤丛枝菌根真菌群落的影响[J]. 土壤, 2022, 54(4): 733−739 doi: 10.13758/j.cnki.tr.2022.04.010XUE Z Z, FENG T Y, WANG C, et al. Effects of land-use patterns on arbuscular mycorrhizal fungi community in acidic red soil[J]. Soils, 2022, 54(4): 733−739 doi: 10.13758/j.cnki.tr.2022.04.010 [31] 彭岳林, 杨敏娜, 蔡晓布. 西藏高原针茅草地土壤因子对丛枝菌根真菌物种多样性的影响[J]. 应用生态学报, 2010, 21(5): 1258−1263 doi: 10.13287/j.1001-9332.2010.0192PENG Y L, YANG M N, CAI X B. Influence of soil factors on species diversity of arbuscular mycorrhizal (AM) fungi in Stipa steppe of Tibet Plateau[J]. Chinese Journal of Applied Ecology, 2010, 21(5): 1258−1263 doi: 10.13287/j.1001-9332.2010.0192 [32] 高威, 王连峰, 贾仲君. 长期不同施肥模式对农田黑土微生物群落构建的影响[J]. 生态与农村环境学报, 2021, 37(11): 1437−1448 doi: 10.19741/j.issn.1673-4831.2020.0807GAO W, WANG L F, JIA Z J. Changes in community assembly of microbiomes in black soil under distinct scenarios of long-term field fertilization[J]. Journal of Ecology and Rural Environment, 2021, 37(11): 1437−1448 doi: 10.19741/j.issn.1673-4831.2020.0807 [33] FENG Y Z, GUO Z Y, ZHONG L H, et al. Balanced fertilization decreases environmental filtering on soil bacterial community assemblage in North China[J]. Frontiers in Microbiology, 2017, 8: 2376 doi: 10.3389/fmicb.2017.02376 [34] ZHANG B G, ZHANG J, LIU Y, et al. Biogeography and ecological processes affecting root-associated bacterial communities in soybean fields across China[J]. Science of the Total Environment, 2018, 627: 20−27 doi: 10.1016/j.scitotenv.2018.01.230 [35] JIAO S, LU Y H. Soil pH and temperature regulate assembly processes of abundant and rare bacterial communities in agricultural ecosystems[J]. Environmental Microbiology, 2020, 22(3): 1052−1065 doi: 10.1111/1462-2920.14815 [36] KNELMAN J E, NEMERGUT D R. Changes in community assembly may shift the relationship between biodiversity and ecosystem function[J]. Frontiers in Microbiology, 2014, 5: 424 [37] GRAHAM E, STEGEN J. Dispersal-based microbial community assembly decreases biogeochemical function[J]. Processes, 2017, 5(4): 65 doi: 10.3390/pr5040065 [38] OLIVERES S, VAN DER PLAS F, MANNING P, et al. Biodiversity at multiple trophic levels is needed for ecosystem multifunctionality[J]. Nature, 2016, 536(7617): 456−459 doi: 10.1038/nature19092 -

 下载:
下载:






 点击查看大图
点击查看大图

计量
- 文章访问数: 46
- HTML全文浏览量: 13
- PDF下载量: 6
- 被引次数: 0
