-
中国农业碳排放效率测度、空间溢出与影响因素
吴昊玥, 黄瀚蛟, 何宇, 陈文宽
-
耕作与秸秆还田方式对碳氮在土壤团聚体中分布的影响
张玉铭, 胡春胜, 陈素英, 王玉英, 李晓欣, 董文旭, 刘秀萍, 裴林, 张惠
-
政策激励、生态认知与农户有机肥施用行为——基于有调节的中介效应模型
桑贤策, 罗小锋, 黄炎忠, 唐林
-
基于InVEST和CA-Markov模型的黄河流域碳储量时空变化研究
杨洁, 谢保鹏, 张德罡
-
秸秆还田与施肥方式对稻麦轮作土壤细菌和真菌群落结构与多样性的影响
张翰林, 白娜玲, 郑宪清, 李双喜, 张娟琴, 张海韵, 周胜, 孙会峰, 吕卫光

官方微信公众号


网购期刊二维码
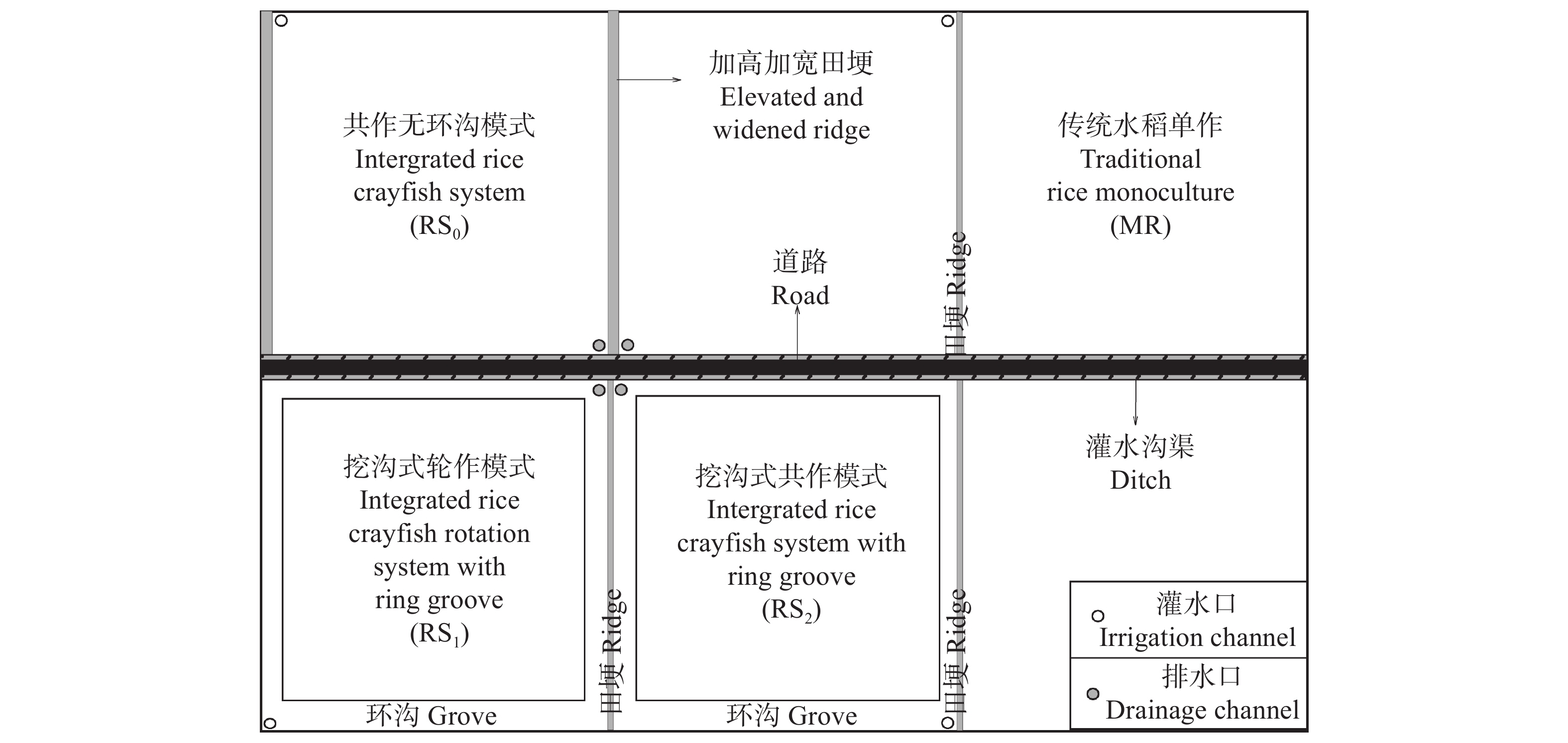
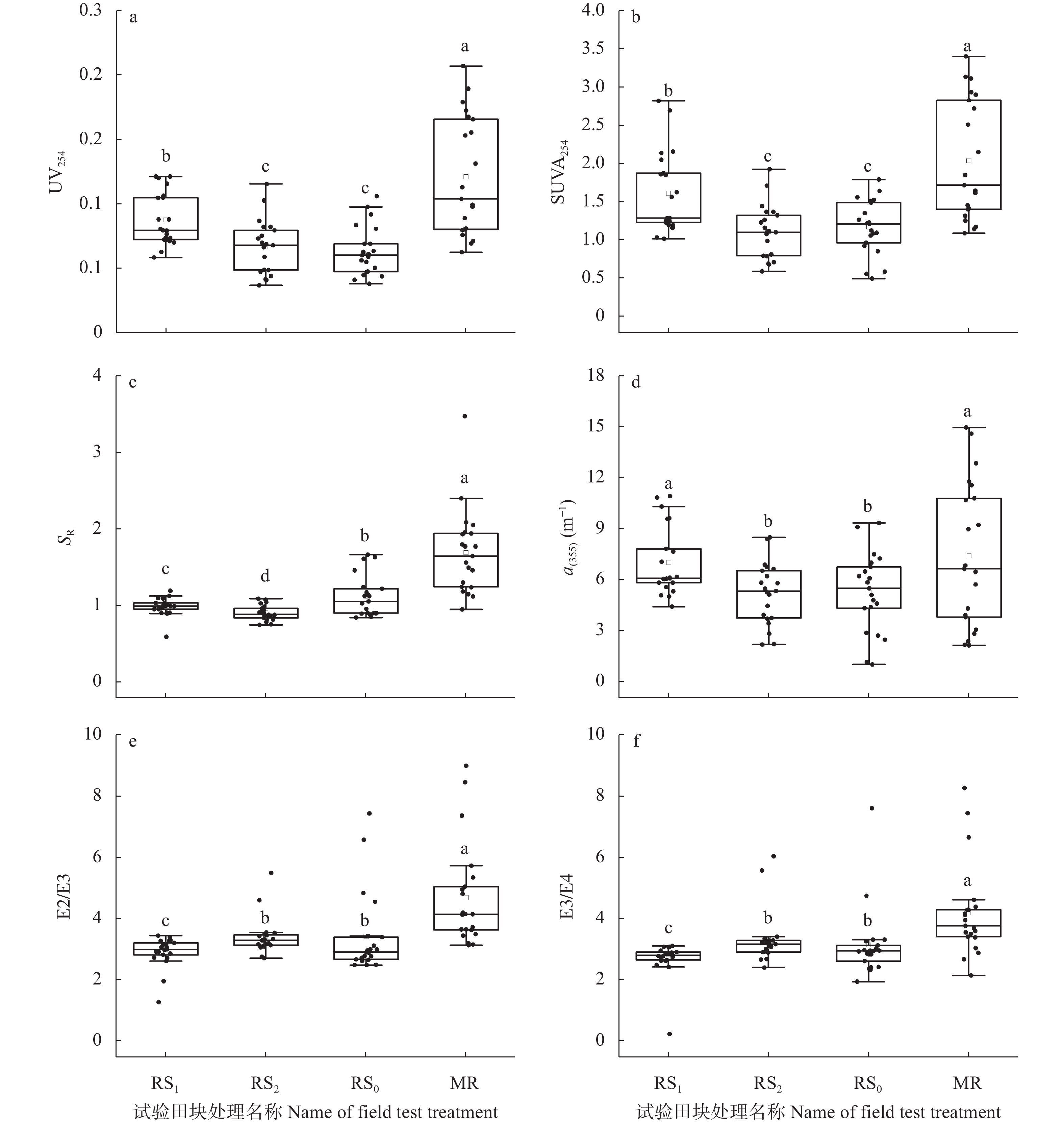
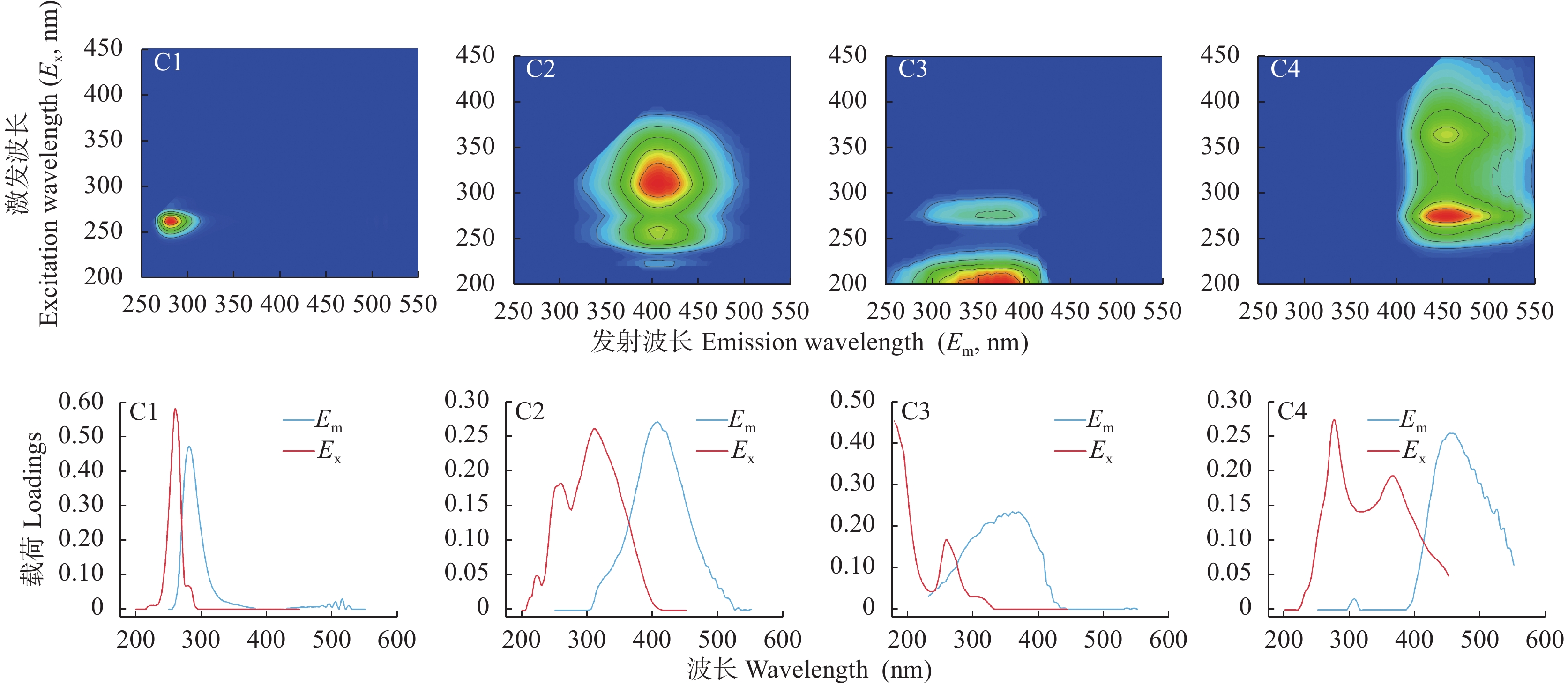
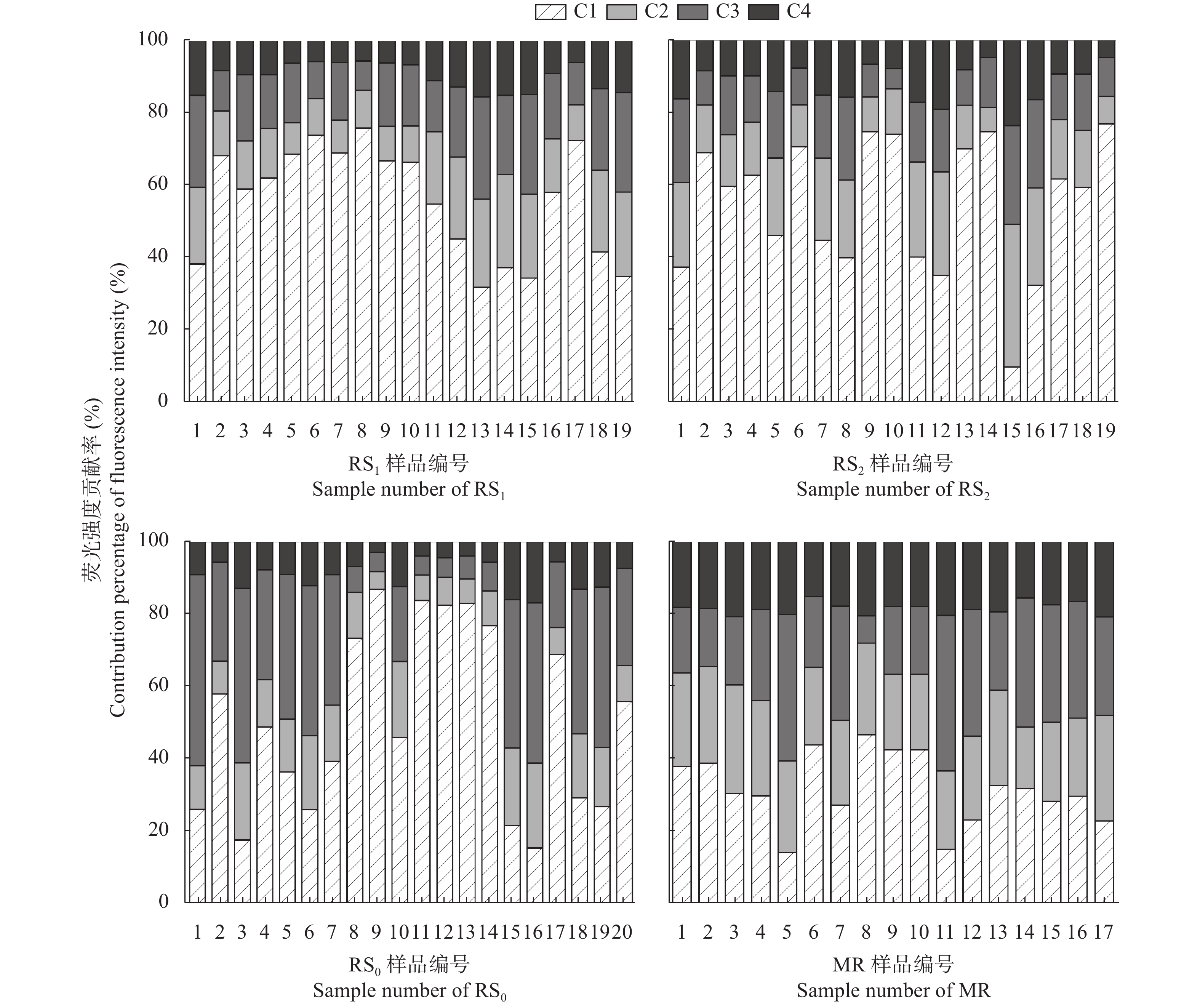
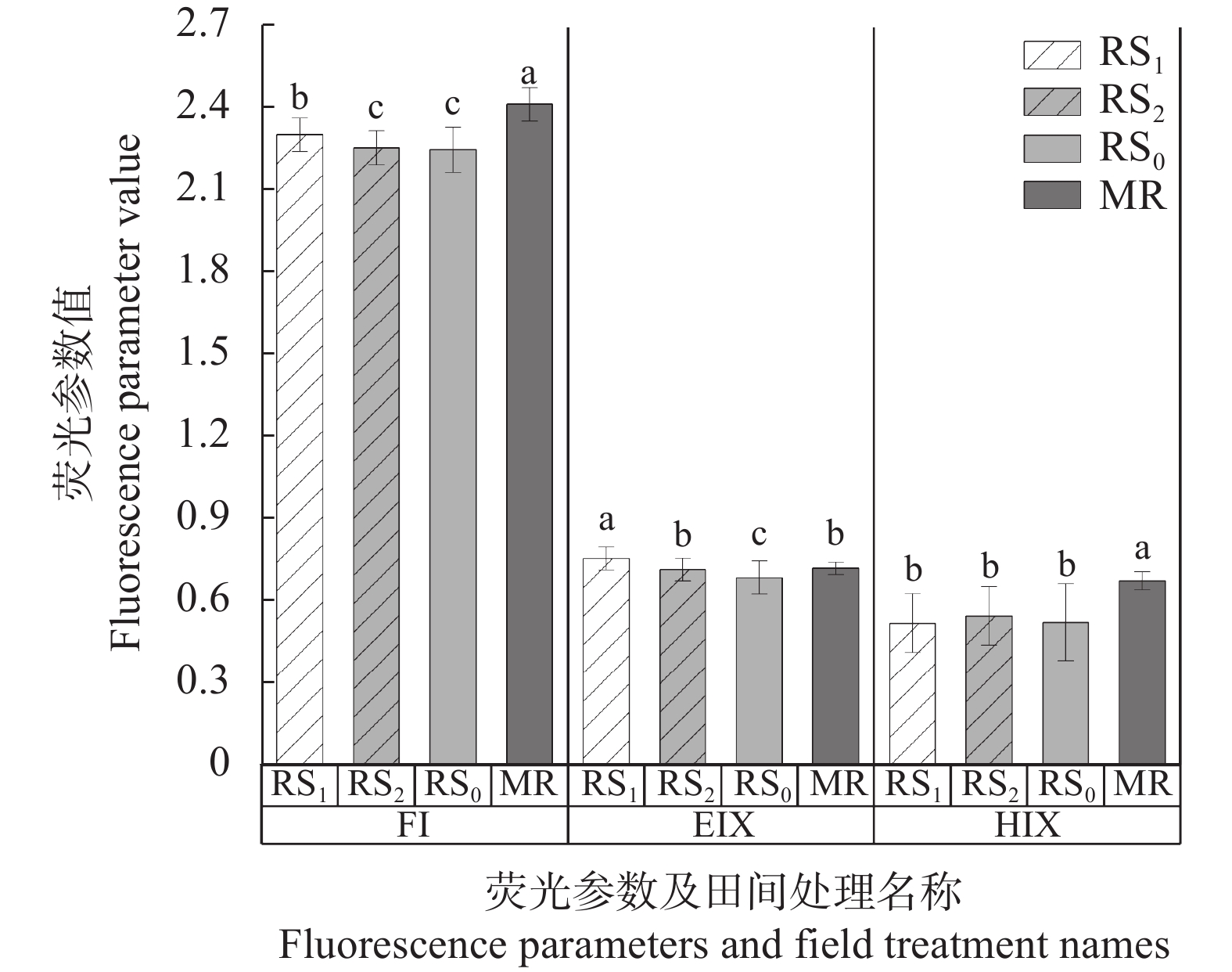
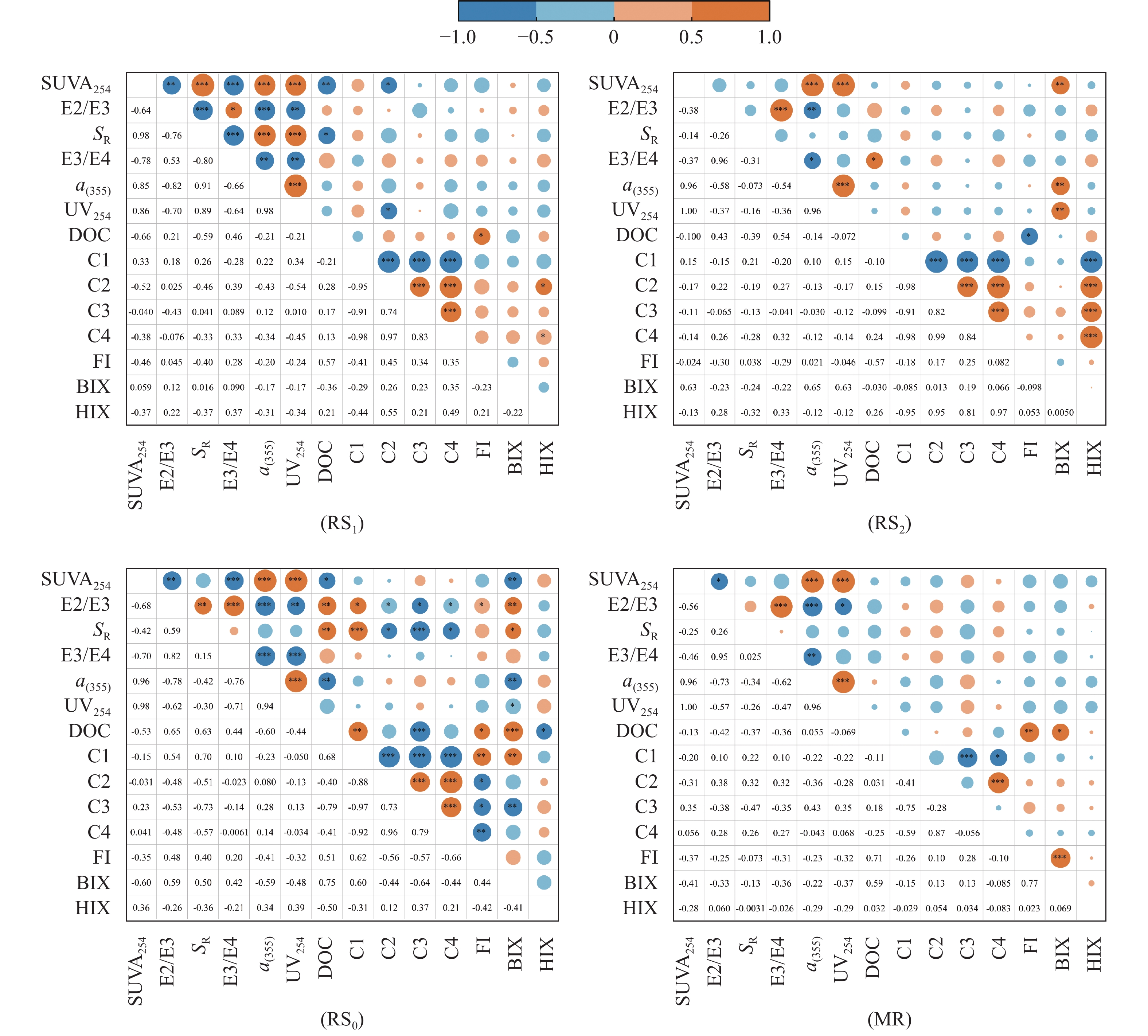
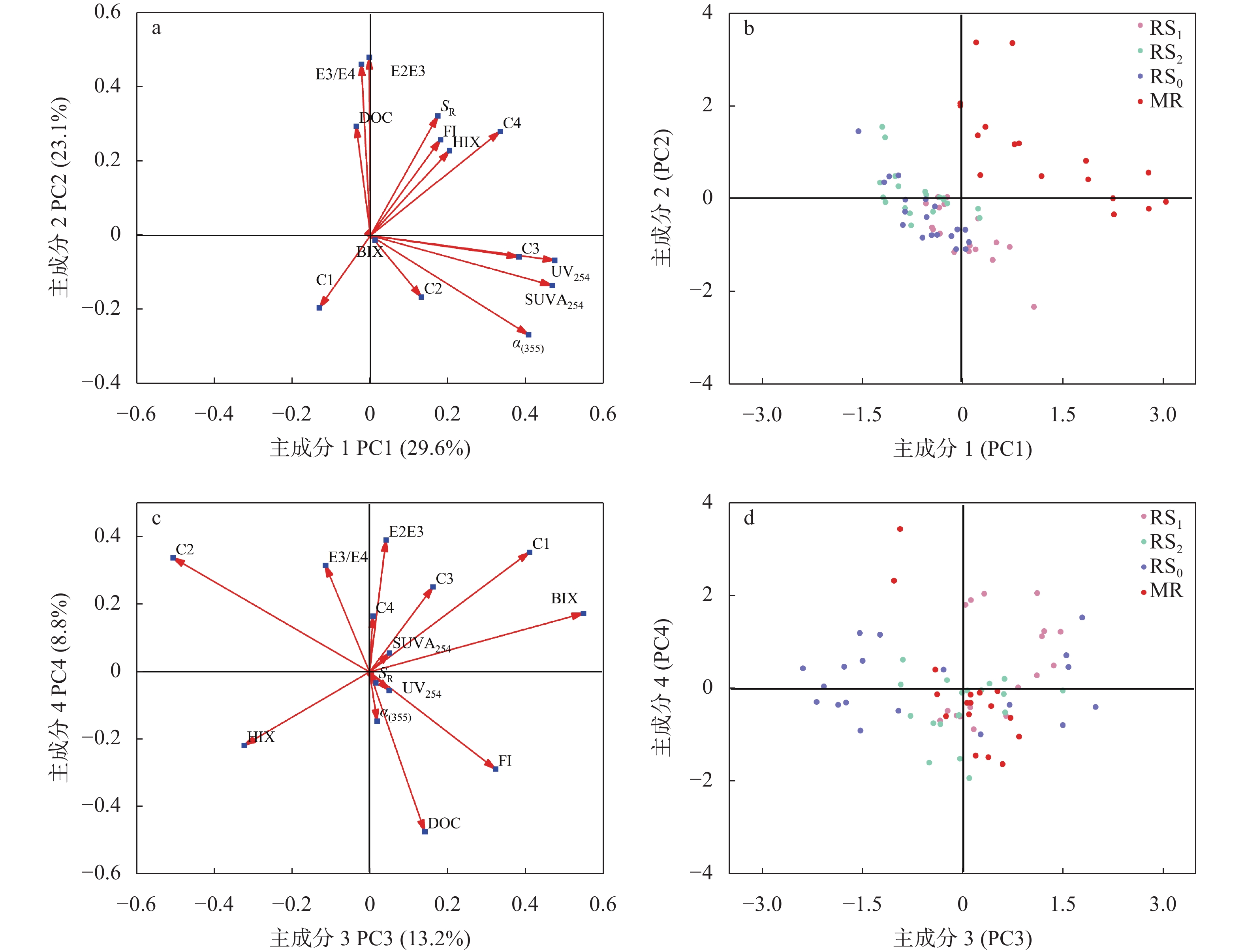
稻虾种养的农业活动显著改变了田间生物化学因子, 土壤溶解性有机质(DOM)是其中之一。本研究利用紫外-可见吸收光和三维荧光光谱技术, 耦合平行因子分析, 对有环沟式稻虾轮作和共作、无环沟式稻虾共作和传统稻作4种稻虾种养模式土壤DOM的特征进行了研究。结果显示, 不同稻虾种养模式土壤DOM紫外吸光度和比吸收系数均低于传统稻作, 光谱斜率比(SR)均值为0.9~2.0; 254 nm和365 nm处紫外吸光度的比值(E2/E3)在无环沟稻虾共作模式中最小, 水稻单作中E2/E3值相对于稻虾种养模式大; 300 nm和400 nm处紫外吸光度的比值(E3/E4)在稻虾种养模式均<3.5, 水稻单作>3.5。4种模式的荧光指数>1.9, 腐殖化指数<4, 自生源指数为0.6~0.7。不同模式均解析出2个类蛋白质组分(类络氨酸、类蛋白物质)和2个类腐殖质组分(腐殖酸、类腐殖质), 与生物源密切相关的类络氨酸组分占比较高且与其他组分均呈负相关, 其余组分相互呈正相关。上述结果中, 紫外-可见吸收光谱参数表明稻虾种养降低了土壤DOM的腐殖化程度、芳香性和物质组成, 稻虾种养土壤腐殖质类DOM腐殖酸为主, 水稻单作以富里酸为主; 4种模式的土壤DOM既有内源也有外源, 其中无环沟稻虾共作模式土壤DOM分子量最大, 且有一定外源性特征, 说明土壤保肥能力较好, 在4种模式中综合效益最佳; 三维荧光光谱分析表明不同模式下土壤DOM的物质组成无明显差异, 研究认为这与农田管理措施的间接影响等有关, 占比较高的络氨酸组分来源不同于其他组分, 可能与微生物对DOM或其他物质的分解转化有关, 而腐殖酸、类蛋白物质和类腐殖质组分很可能有高度的同源性; 不同种养模式之间的淹水差异通过影响溶解性有机碳(DOC)和DOM的释放, 从而影响种养模式之间DOM光谱特征。
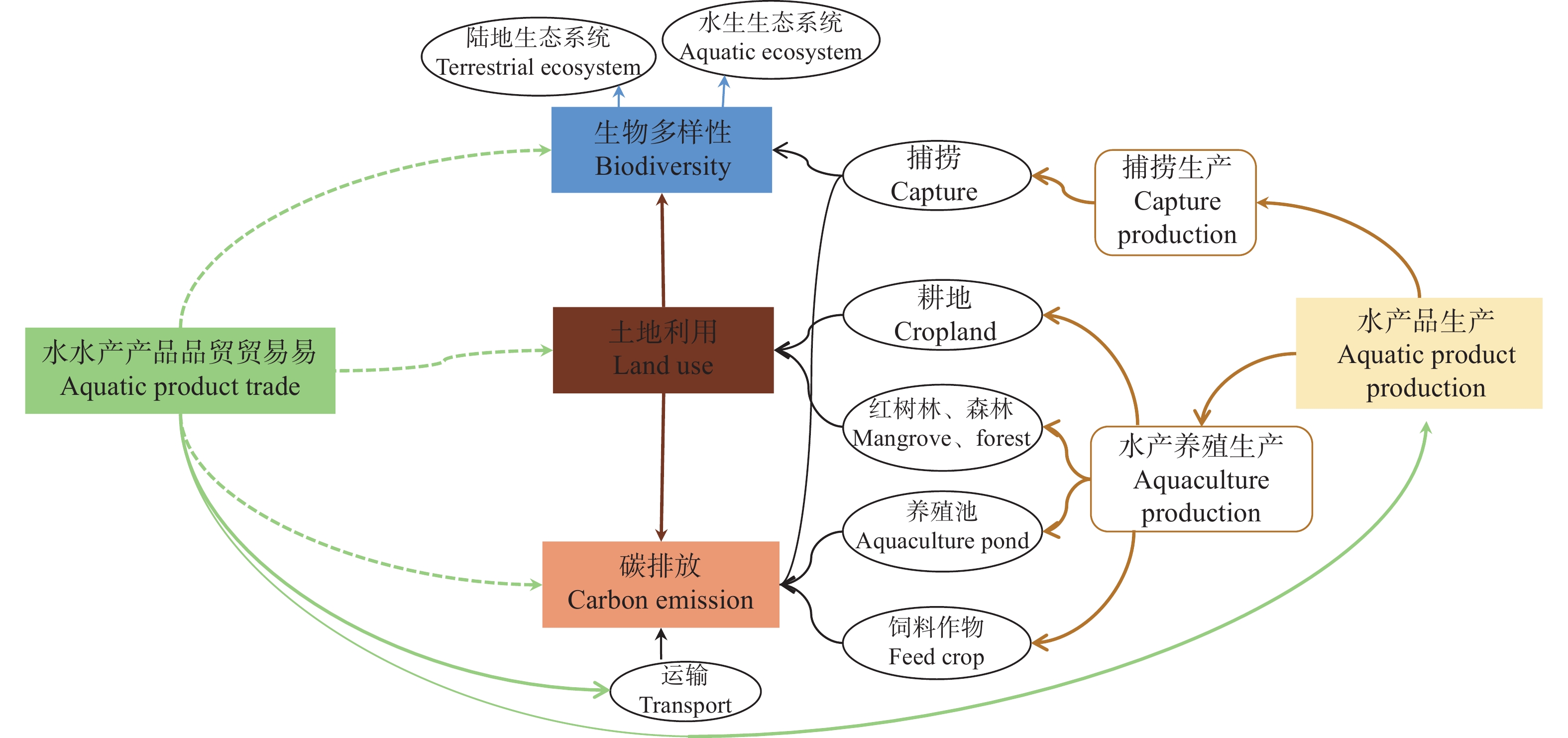
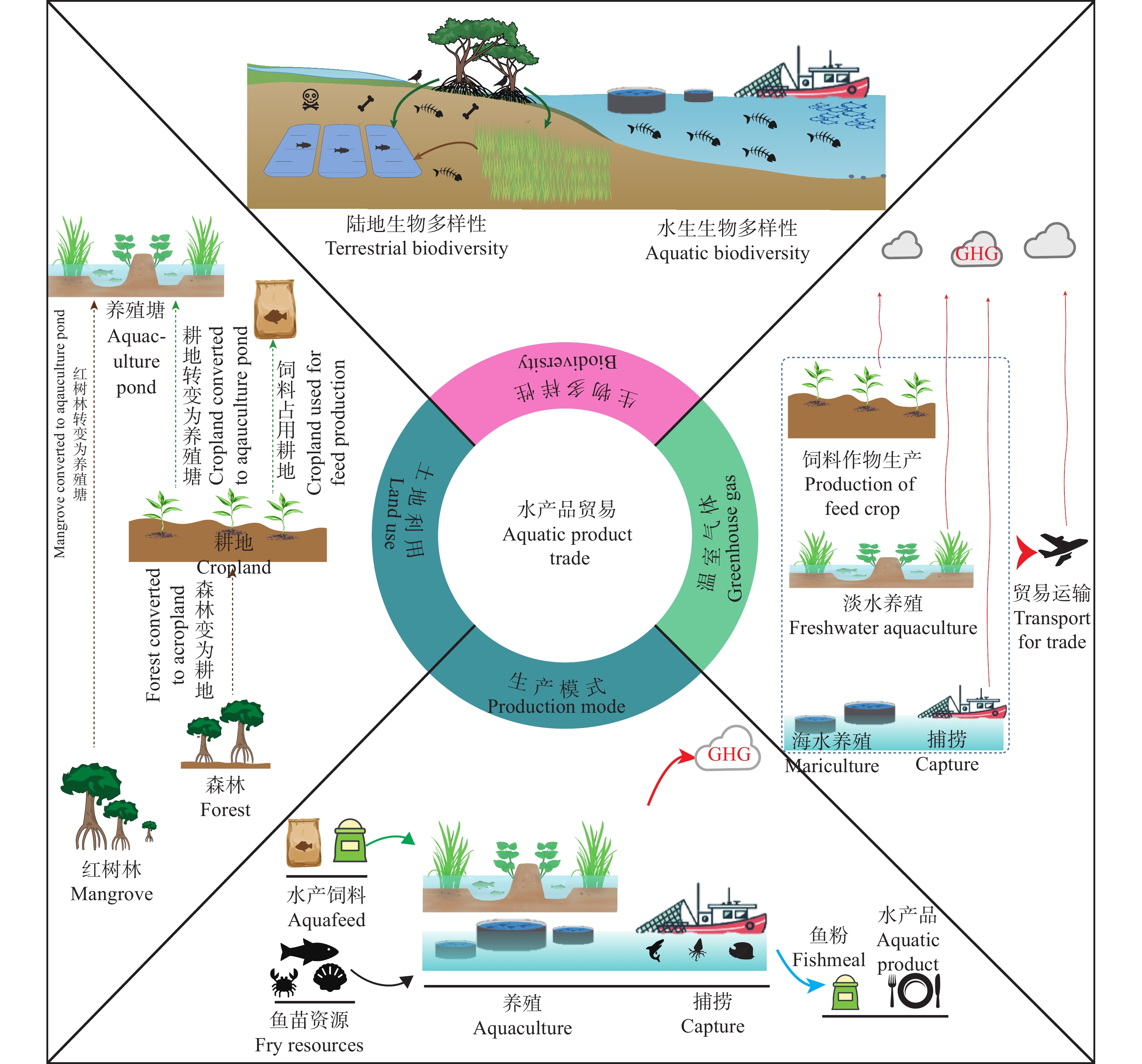
目前全球约37%的水产品进入国际贸易而非本地消费, 因此水产品贸易与全球资源和环境可持续性的联系日益密切。然而, 现有研究多集中于分析水产品替代畜禽产品导致的资源环境代价变化, 而针对水产品贸易与“资源-环境-生物多样性”影响的研究还较缺乏。本文利用环境足迹和全生命周期相结合的方法, 综述了水产品的贸易量、贸易品种与贸易国家的变化规律, 并依据贸易与资源及环境的关系, 分析了水产品贸易对土地利用、碳排放和生物多样性的影响。结果发现, 2020年水产品出口量较1976年增长了5倍, 且贸易增速呈现出“先快后稳”的趋势。水产品贸易国由欧洲南部扩大至全球范围。捕捞水产品, 如沙丁鱼、鳕鱼和金枪鱼是主要的贸易品种; 养殖水产品在总水产贸易产品中的比重快速增加, 由1976年的5%增加至2020年的25%。水产品贸易中养殖产品的增加影响全球土地利用变化、虚拟温室气体排放, 以及水生和陆地生态系统生物多样性。因此, 未来若要实现全球水产品可持续生产和消费, 需发达国家和发展中国家间共享水产品先进生产技术、优化贸易结构、调整贸易品种。例如, 在生产端优化养殖结构、技术和产业链, 在需求端减少对高资源环境代价品种的消费, 在贸易端限制高资源环境代价品种的交易。

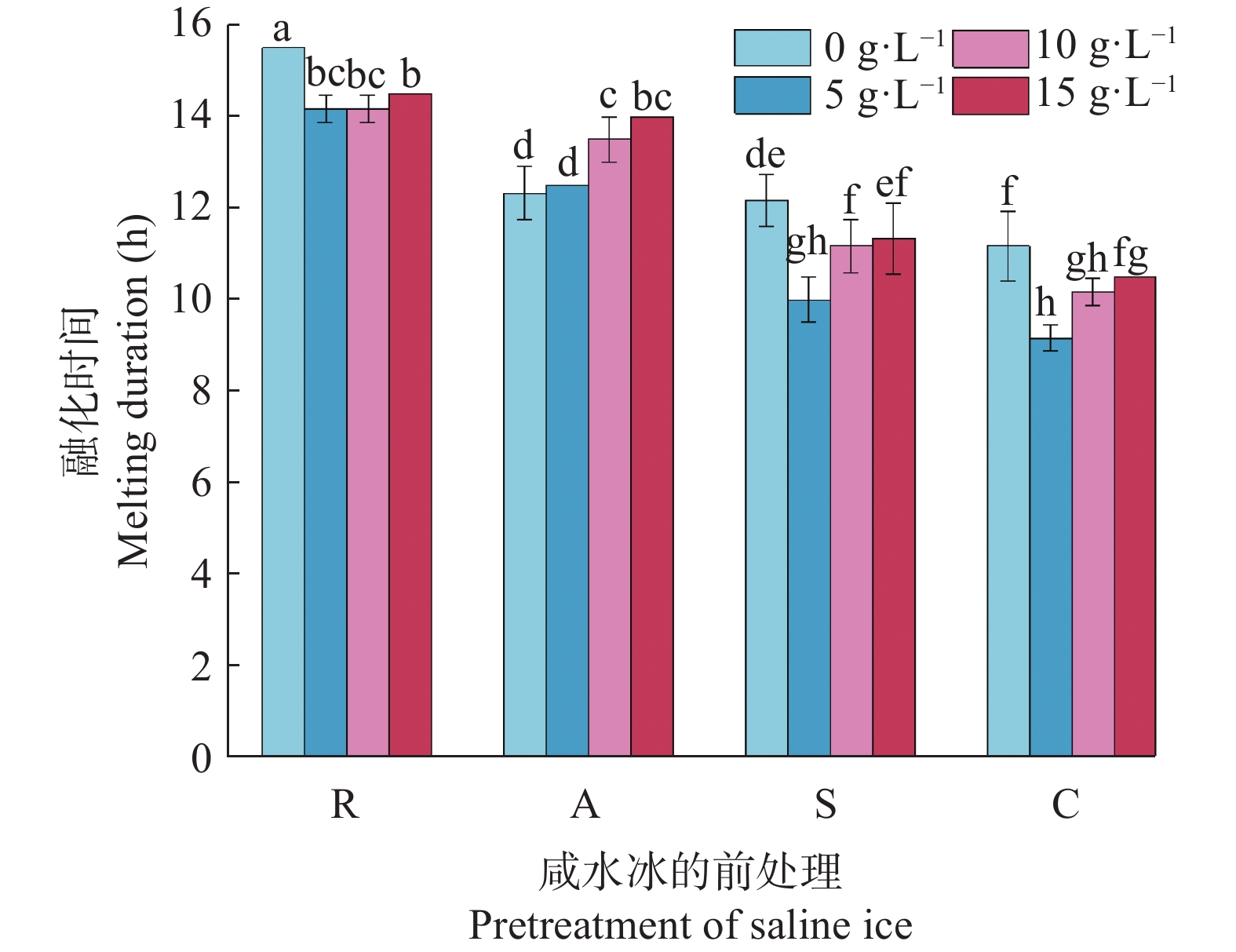
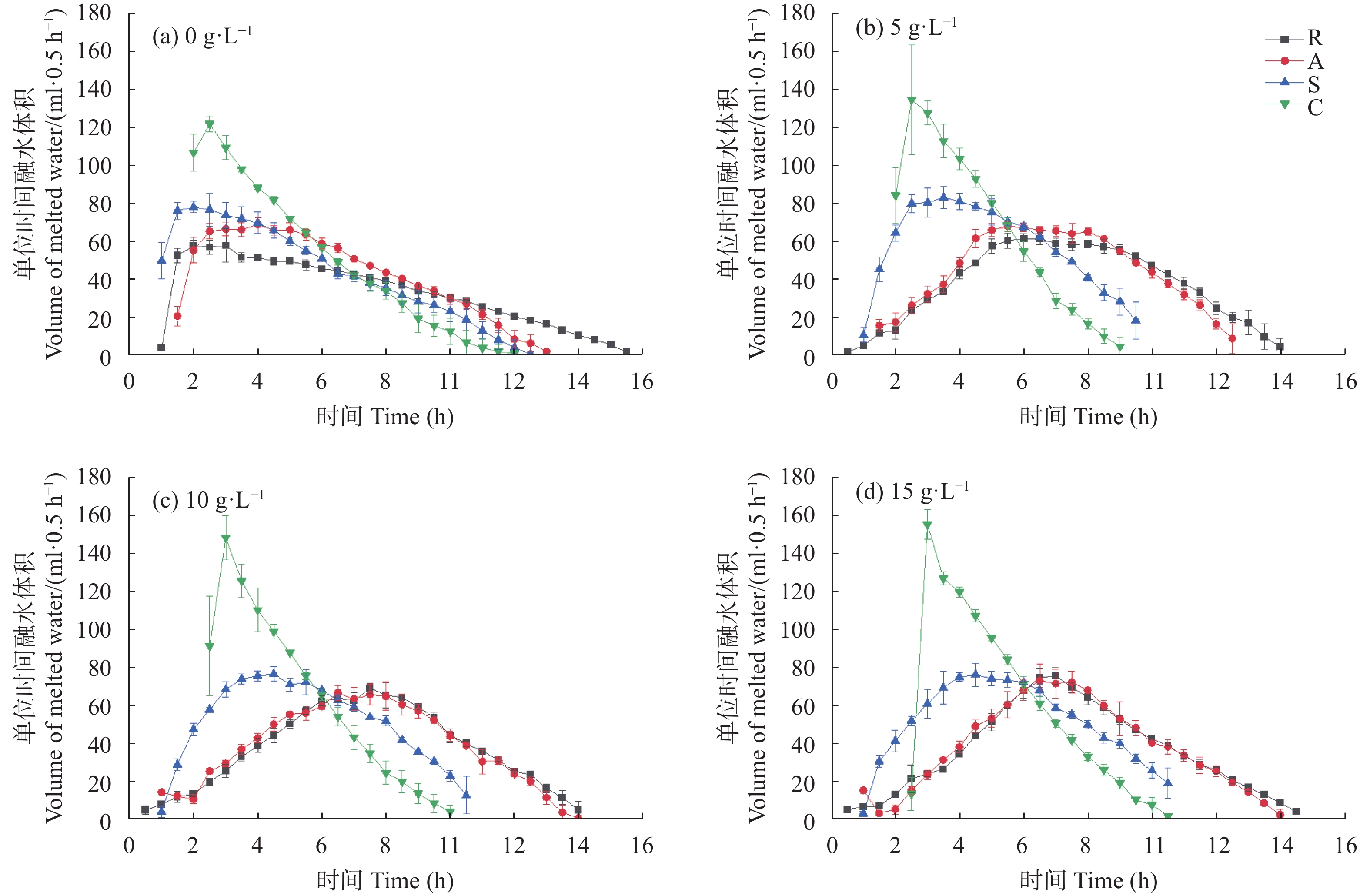
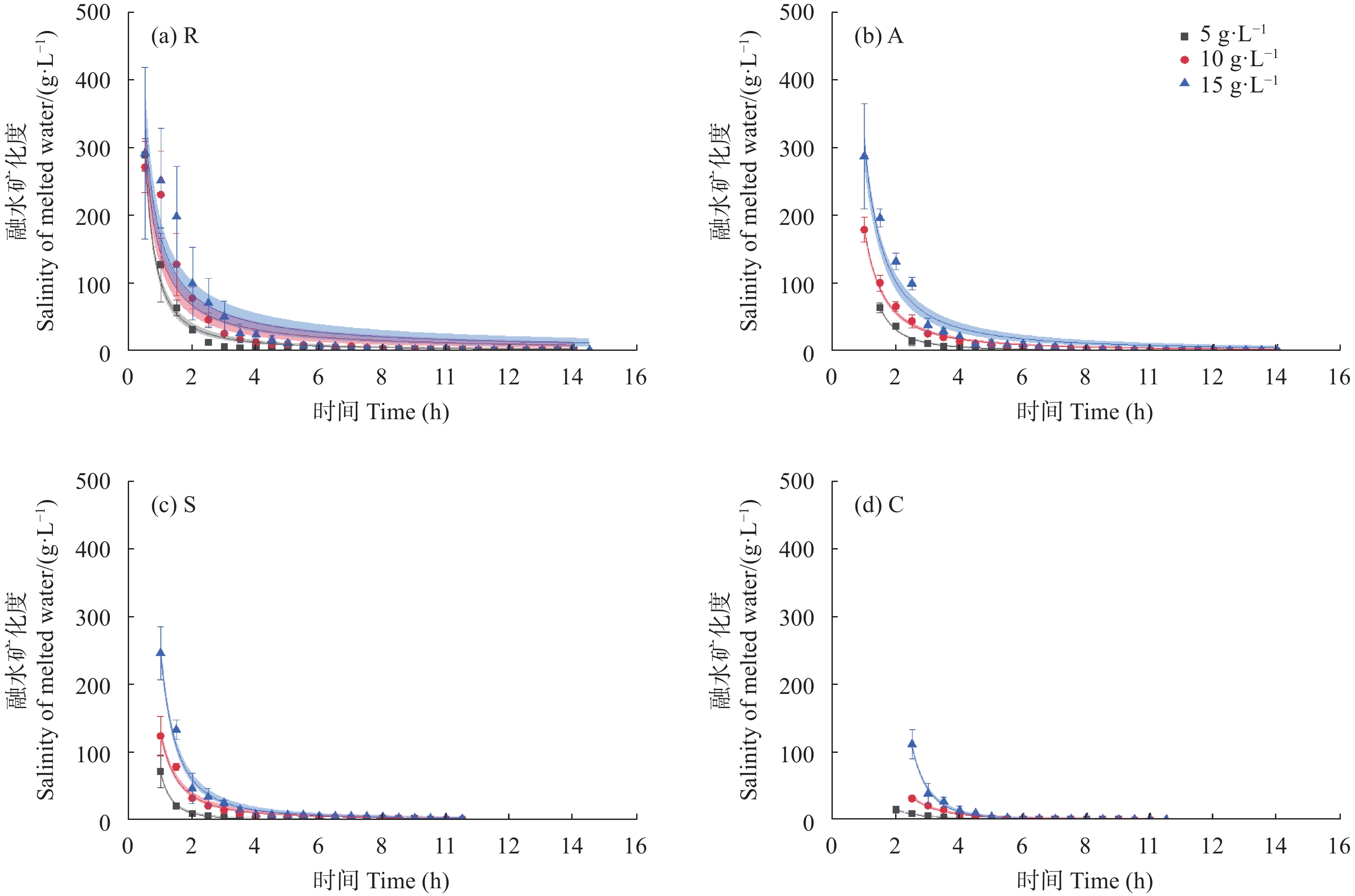
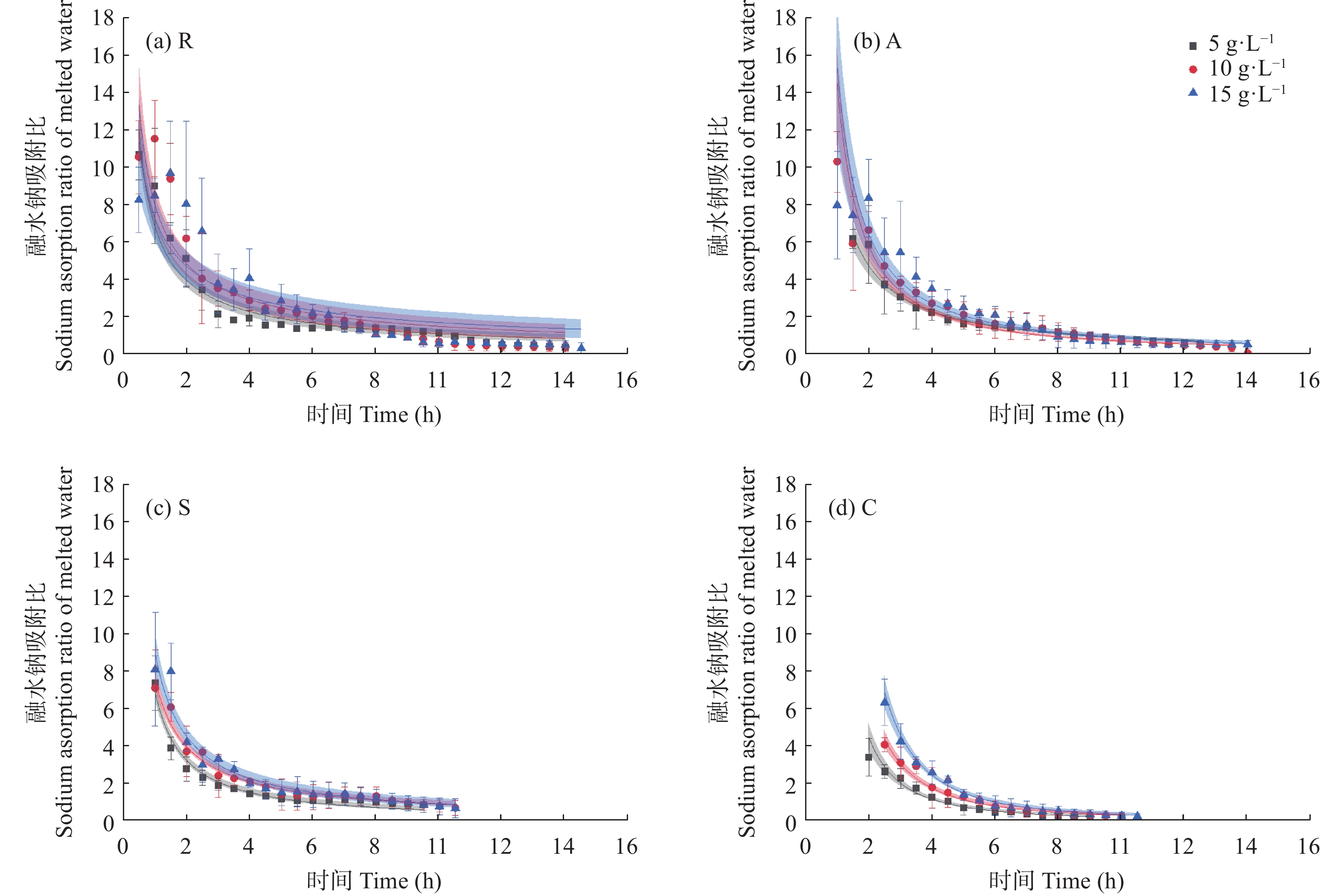
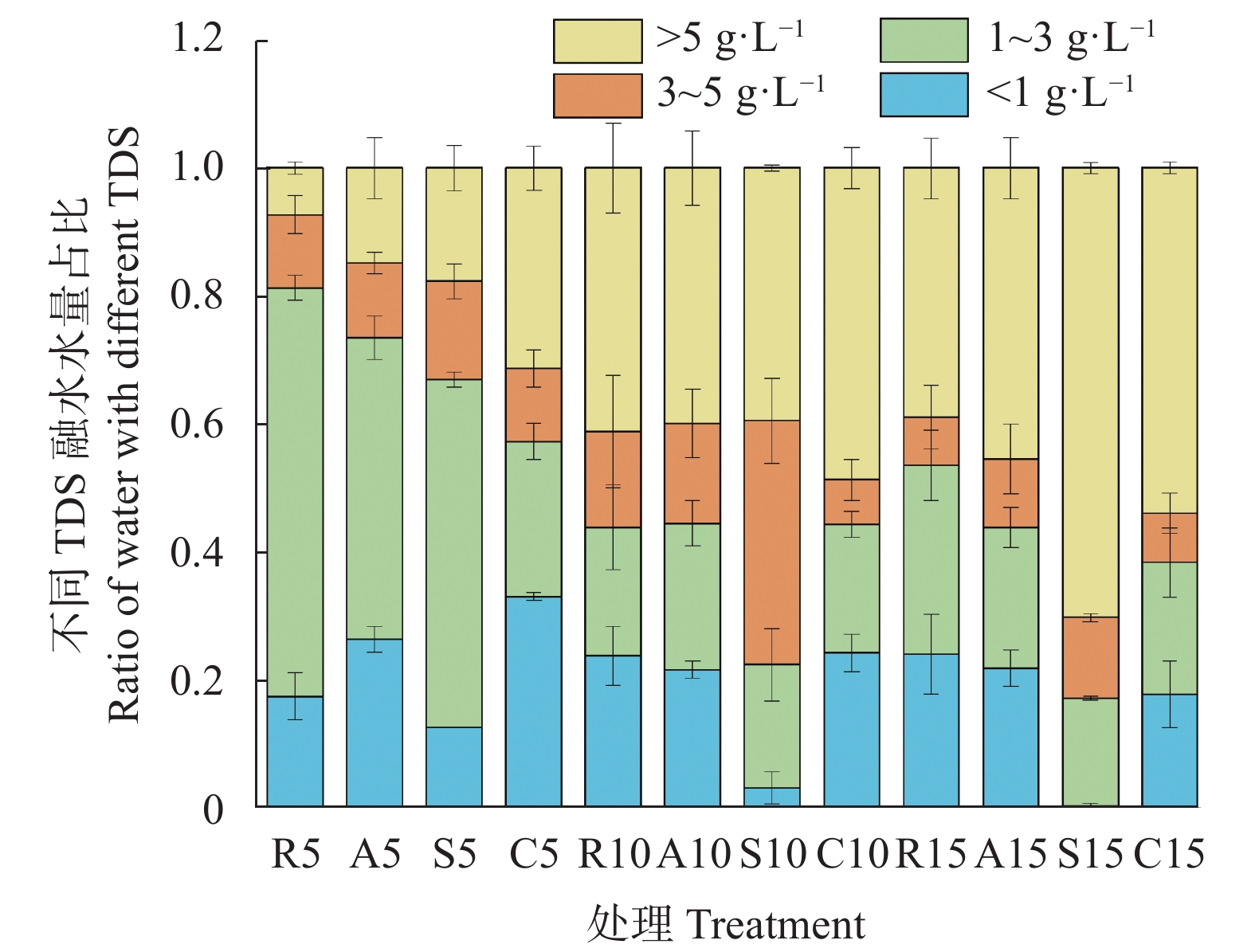
咸水结冰灌溉对重度盐碱地改良, 缓解淡水资源短缺具有重要现实意义。咸水冰融化过程中水盐分离的实质是盐分的再分配过程。为厘清前处理对咸水冰融化过程中水质及水量的影响, 本研究设计了曝气冰(A)、堆积冰(S)、破碎冰(C)和常规冰(R)4种前处理, 及4个初始矿化度0、5 g∙L−1、10 g∙L−1、15 g∙L−1, 对比分析融水过程中水量和水质的动态变化。结果表明, 曝气、堆积和破碎处理相对常规冰都能显著减少融化时间, 各处理融化时间为破碎冰<堆积冰<曝气冰<常规冰; 单位时间融出水量均呈现先快速升高, 后缓慢降低的趋势, 峰值水量为常规冰<曝气冰<堆积冰<破碎冰。随着融化过程的进行, 融水矿化度快速下降, 之后逐渐平缓; 其中, 破碎冰初始和结束时融水矿化度和钠吸附比均显著低于常规冰; 且初始矿化度为5 g∙L−1时破碎冰的淡水(<1 g∙L−1)融出量高达33.26%, 显著高于同矿化度的其他前处理冰体。可见, 前处理措施可以显著改变咸水冰融化过程中的水盐再分配过程, 这为地下咸水资源利用时通过咸水冰前处理提高盐分淋洗能力提供了理论依据。

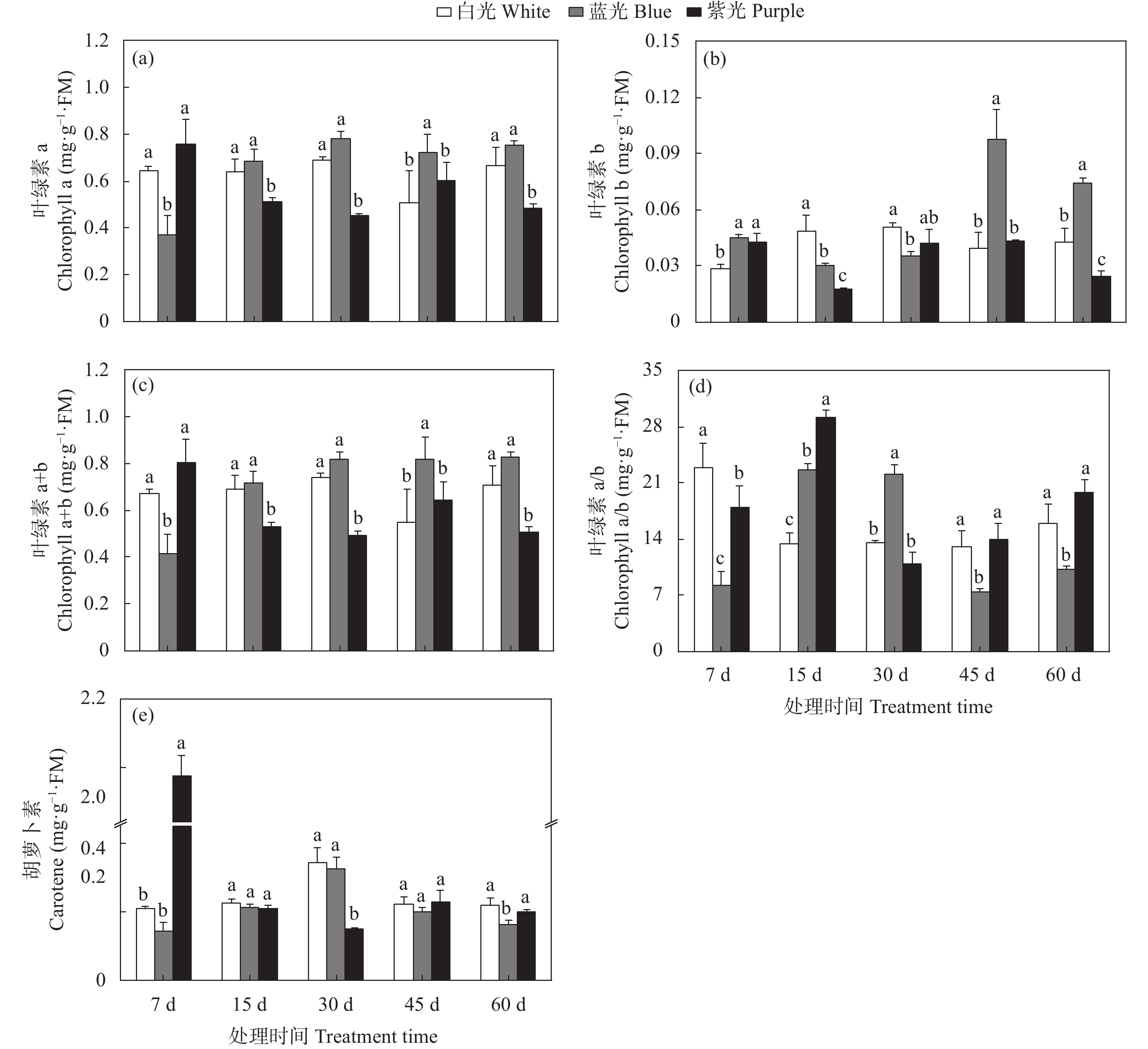
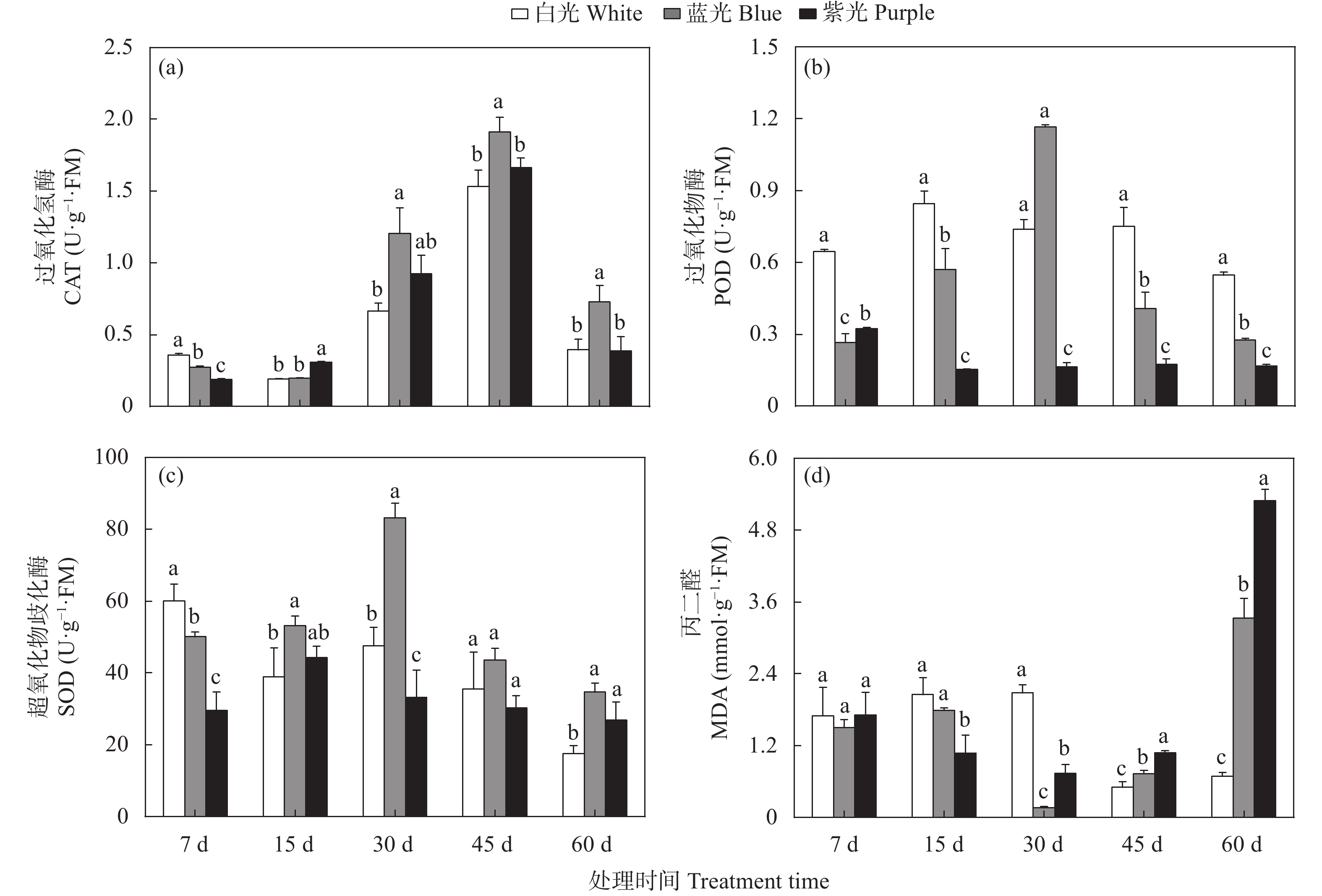
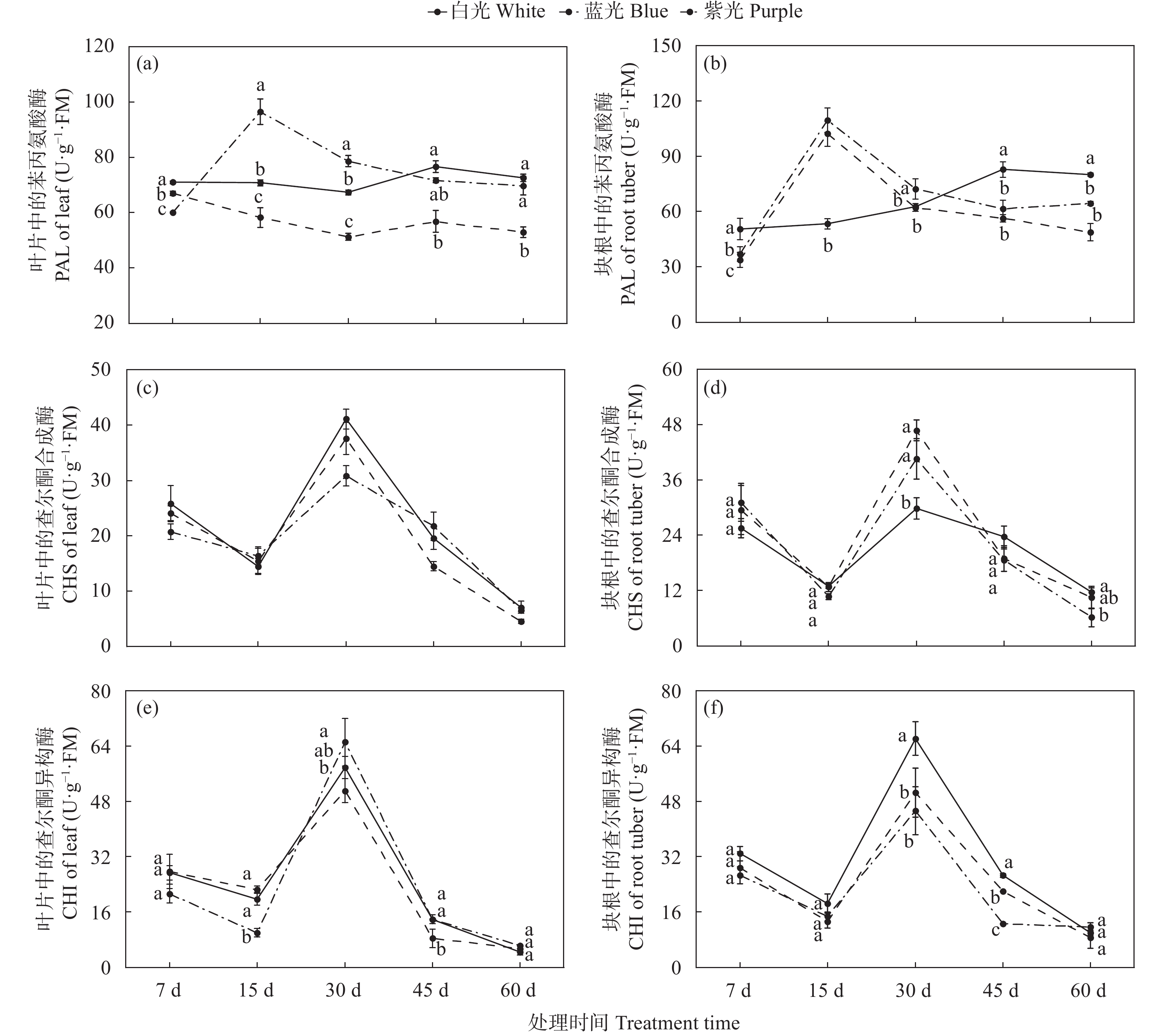
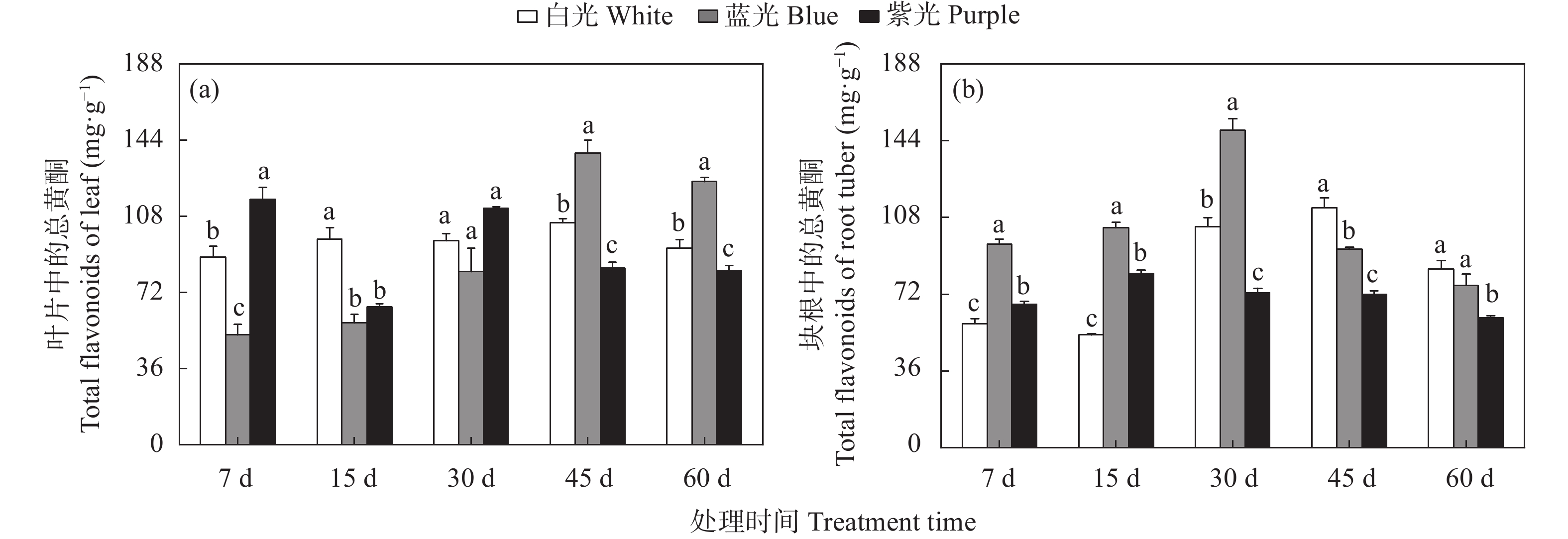
三叶青作为“新浙八味”之一的药材, 具有抗肿瘤、抗病毒和退热等临床效果, 其主要有效成分为黄酮类化合物。三叶青生长在林间荫蔽环境, 其活性成分合成、生长发育、生理变化等均会受到林下短波光调控。为明晰短波光对三叶青品质及生理变化的调控规律, 本文选择了蓝光和紫光单色光作为处理光源, 以白光作为对照, 处理7 d、15 d、30 d、45 d和60 d, 测定三叶青总黄酮含量、黄酮代谢合成酶活性、生理生化指标(光合色素、抗逆酶活性和丙二醛含量、初生代谢可溶物), 以期探讨三叶青黄酮代谢和生理生化变化对短波光条件的时空响应及规律。结果表明: 持续的单色短波光处理, 使三叶青处于轻度胁迫状态, 蓝光处理后三叶青中总黄酮含量最高, 其中叶片含量为137.75 mg∙g−1, 块根中为149.00 mg∙g−1; 紫光处理使叶片和块根中的PAL酶活性(96.50 U∙g−1FW、109.47 U∙g−1FW)和叶片中CHI酶活性(65.17 U∙g−1 FW)均达最高, 蓝光处理块根中的CHS酶活性最高, 为46.76 U∙g−1 FW。此外, 光合色素和抗逆能力和可溶性物质对短波光的响应度有差异, 与白光相比, 蓝光和紫光均可促进渗透调节物质的积累。本研究的结果可为光调控三叶青品质提供理论基础和实践指导。

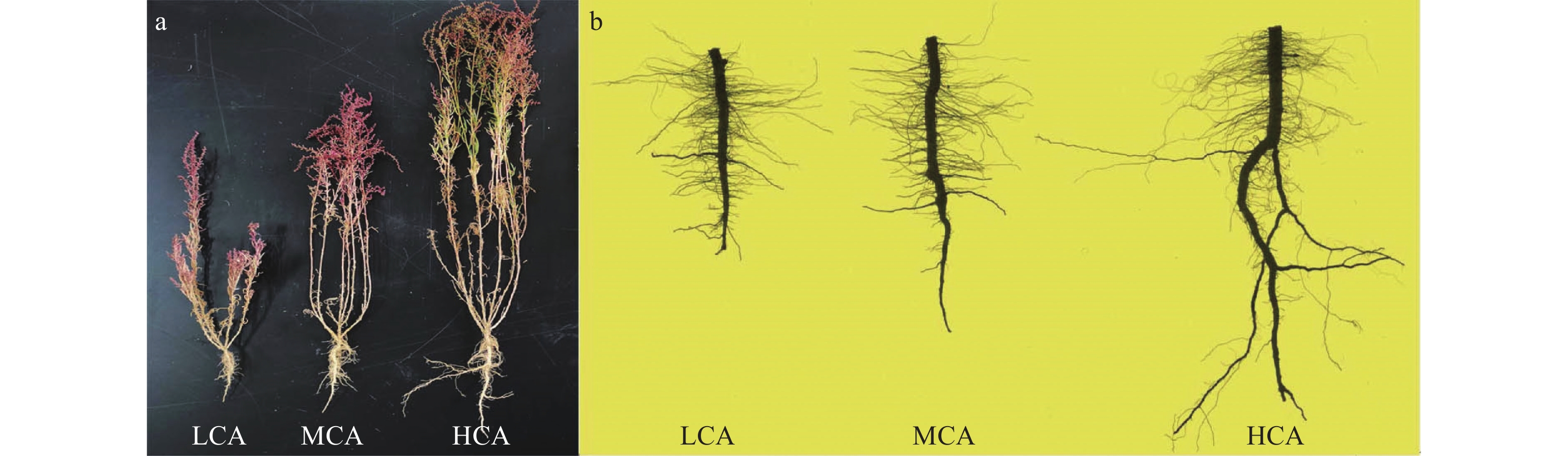
为探究不同覆盖度下盐地碱蓬的生长特征及土壤呼吸速率变化规律, 以黄河三角洲滨海滩涂典型的盐地碱蓬群落为研究对象, 选取滩涂裸地区、低覆盖区、中覆盖区和高覆盖区4种样地, 研究了不同覆盖度下盐地碱蓬的植株生长、根系分布及土壤呼吸速率变化间的差异。结果表明: 盐地碱蓬不同覆盖区土壤的理化性质和植被生长状况差异明显, 与滩涂裸地相比, 盐地碱蓬不同覆盖区土壤含盐量和容重降低, 土壤孔隙度和养分增加。盐地碱蓬的生长指标与覆盖度呈正相关, 生物量、株高及分支数均随覆盖度的提高显著增加(P<0.05)。其中, 盐地碱蓬地下部分生物量主要集中在0~20 cm表层土壤中, 呈现浅层化分布; 且均以2~5 mm粗度根系为主, 分别占低、中和高覆盖区地下生物量的72.53%、59.72%和39.30%。细根的根长、表面积、根尖数、分支数和交叉数均随覆盖度的提高而逐步增大, 且不同覆盖区之间差异显著(P<0.05)。不同覆盖区内土壤呼吸速率表现为高覆盖区>中覆盖区>低覆盖区>裸地区, 并具有明显的日变化, 呈现出低-高-低的单峰曲线, 最大值出现在12:00—14:00。相关性分析表明, 土壤含盐量与盐地碱蓬各生长指标呈显著或极显著负相关, 是主要限制因子, 而土壤呼吸速率与植株各生长指标均呈极显著正相关。本研究结果可为黄河三角洲滨海滩涂植被恢复与生态修复提供理论依据。
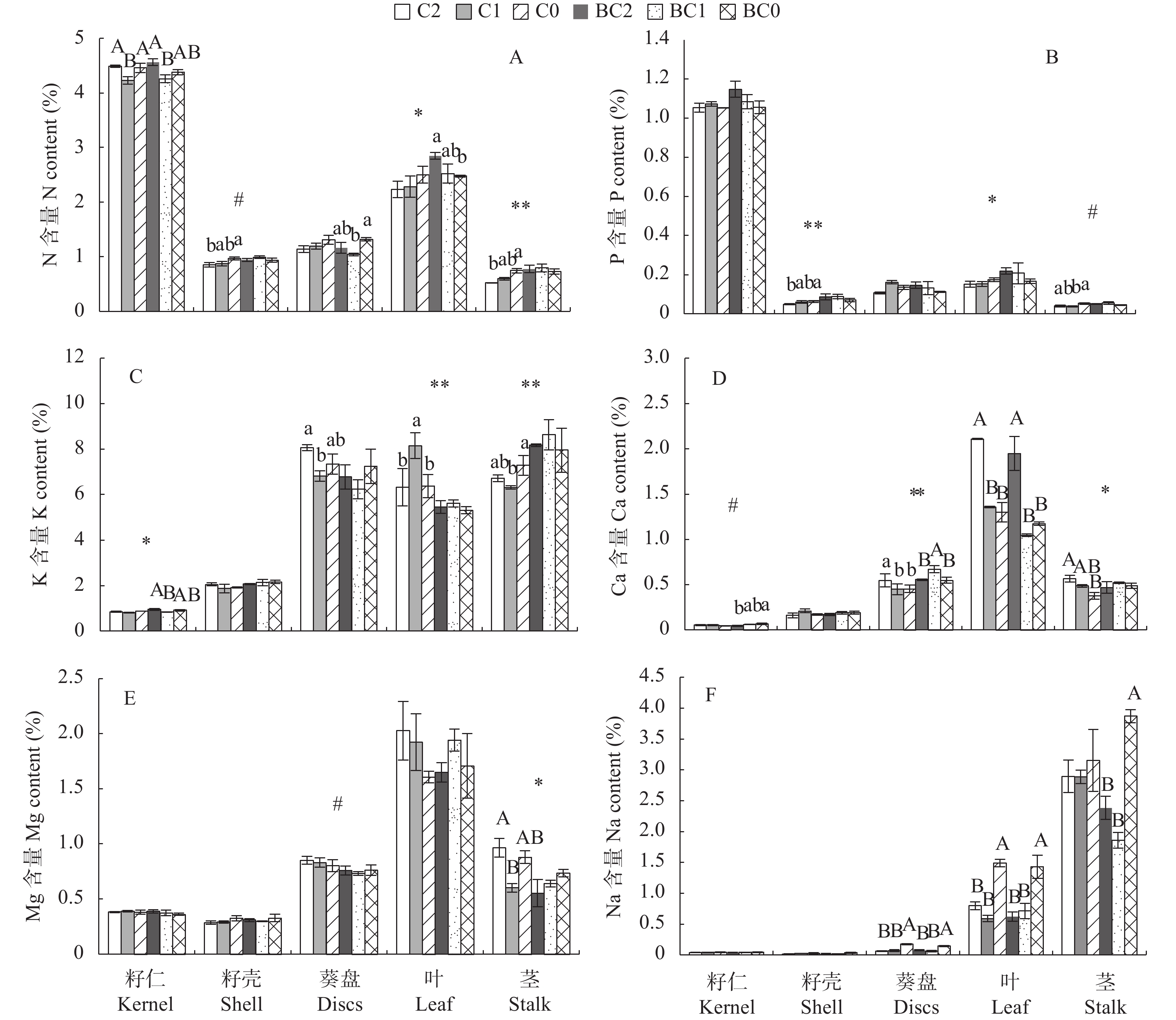
以生物改良、开发利用荒芜重盐碱地为目标, 在华北低平原区滨海荒芜重盐碱区开展了施用生物炭(B)与腐殖酸型土壤调理剂(C)对盐碱地先锋作物棉花和油葵的大田试验。生物炭施用量设2个水平(0 kg∙m−2和1.25 kg∙m−2), 调理剂施用量设3个水平(0 kg∙m−2、0.83 kg∙m−2和1.66 kg∙m−2), 共6个处理。研究结果表明, 生物炭显著抑制油葵早期的植株生长, 对后期生长及籽粒数量和重量无显著影响, 提高了油葵茎、叶和籽壳的N、P含量, 增加了茎、籽壳和籽仁的K含量, 促进叶片K、Ca向籽仁转移; 但施用生物炭抑制油葵吸收Mg, 导致茎秆和葵盘Mg含量显著下降, 对Na的吸收没有显著影响。腐殖酸型土壤调理剂则显著促进油葵茎、叶和葵盘生长, 提高了籽粒产量, 促进P向籽仁转移; 施用1.66 kg∙m−2调理剂处理促进N优先向油葵籽仁转移, 大幅度提升茎、叶和葵盘中的Ca含量, 同时提高茎秆Mg含量; 生物炭与土壤调理剂组合施用可消弱生物炭对油葵生长的抑制作用, 提高油葵对N、P、K、Ca的吸收, 促进N、P、K向籽仁的转移, 降低对Na和Mg的吸收, 增强油葵选择性吸收养分和拒盐能力。本研究还表明, 油葵比棉花更适合作为先锋作物在滨海荒芜重盐碱地种植。研究结果为秸秆等废弃物资源化利用、荒芜重盐碱地开发与生物改良以及耐盐适生先锋作物的选择提供理论依据。
新疆盐碱地类型多且积盐严重, 因此, 探讨不同盐碱胁迫对棉花养分吸收转运和代谢的影响, 揭示棉花对不同盐碱胁迫的耐受机制, 可为新疆不同盐碱地类型棉花栽培提供一定的理论基础。本研究设置对照(CK)、氯化钠(NaCl)盐胁迫(CS)、硫酸钠(Na2SO4)盐胁迫(SS)和碱(NaHCO3+Na2CO3)胁迫(AS) 4个处理, 通过离子组和代谢组学的方法, 探究不同盐碱胁迫对棉花根和叶中营养元素含量以及代谢的影响。结果表明, 盐碱胁迫显著抑制棉花生长, 与CK相比, CS、SS和AS处理的棉花总生物量分别显著(P<0.05, 下同)降低51.7%、47.8%和52.3%, CS处理叶片N含量显著增加, P、K、Ca、Mg和S含量显著降低, 茎中N、P、K、Ca、Mg和S含量均显著降低, 根中N、Ca和Mg含量显著降低, P含量显著增加; SS处理叶中P、Ca和Mg含量显著降低, S含量显著增加, 茎中P、Ca和Mg含量显著降低, S含量显著增加, 根中P和Ca含量显著降低, Mg和S含量显著增加; AS处理叶中P、K、Ca、Mg和S含量显著降低, 茎中N、P、Ca、Mg和S含量显著降低, 根中N、P和S含量显著降低, Mg含量显著增加。CS处理下棉花叶片和根系中分别筛选出7条差异代谢通路, SS处理下棉花叶片和根系中分别筛选出16和29条差异代谢通路, AS处理下棉花叶片和根系中分别筛选出8条和18条差异代谢通路。氯化钠胁迫抑制棉花P、Ca、Mg和S的转运能力, 但是对代谢的影响相对较小; 硫酸钠胁迫下棉花体内积累的S促进了氨基酸类代谢; 碱胁迫抑制了P、K、Ca、Mg和S的转运能力, 根部有机酸代谢增强且有机酸显著积累, 叶片中亚油酸显著积累。
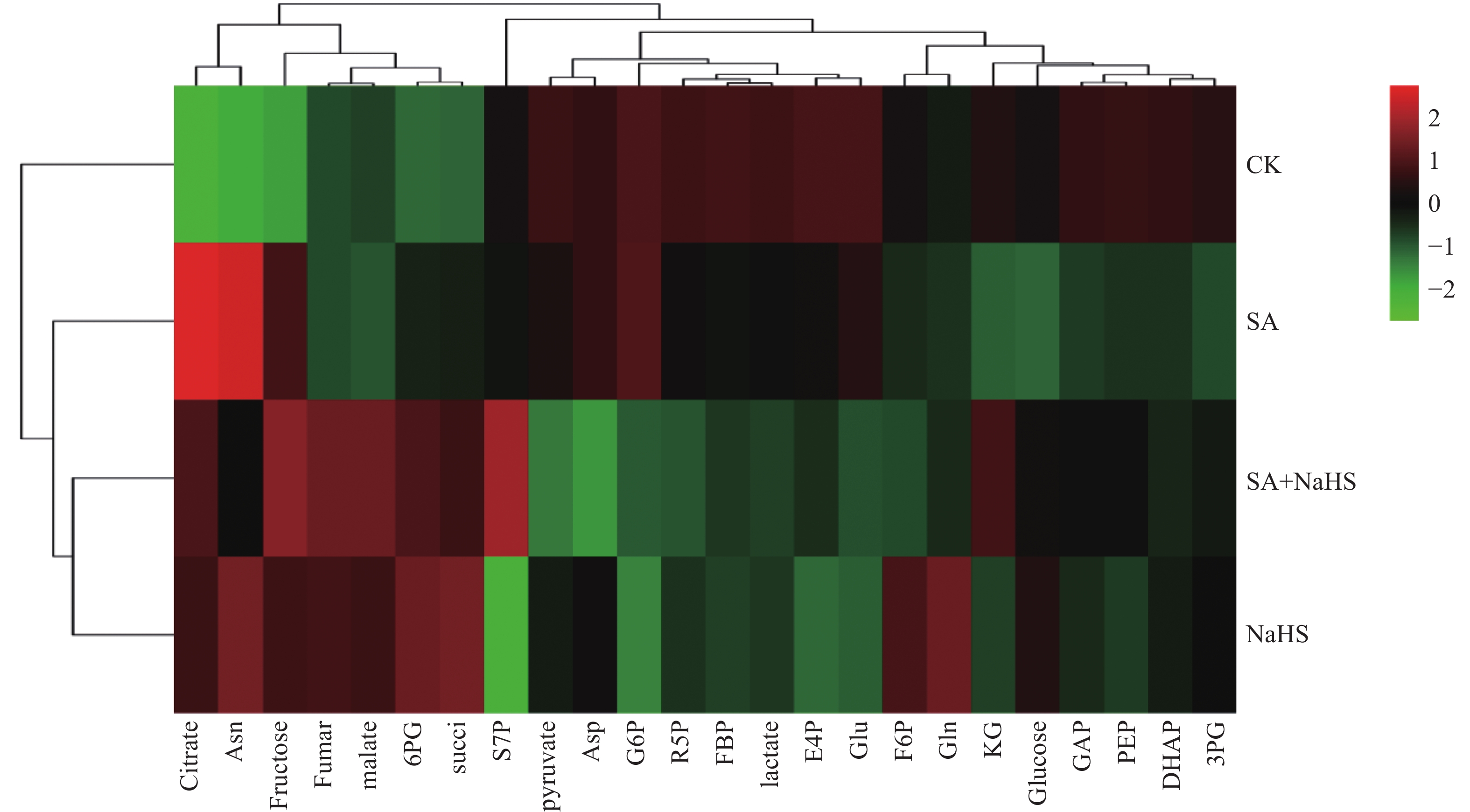
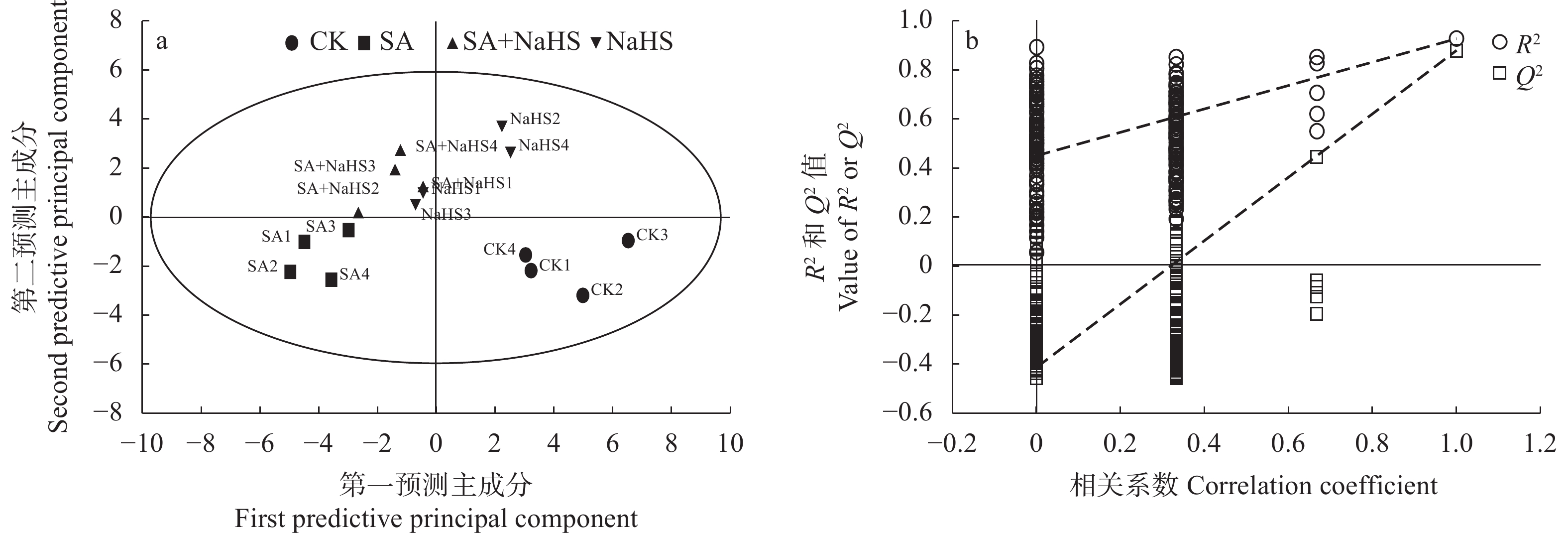
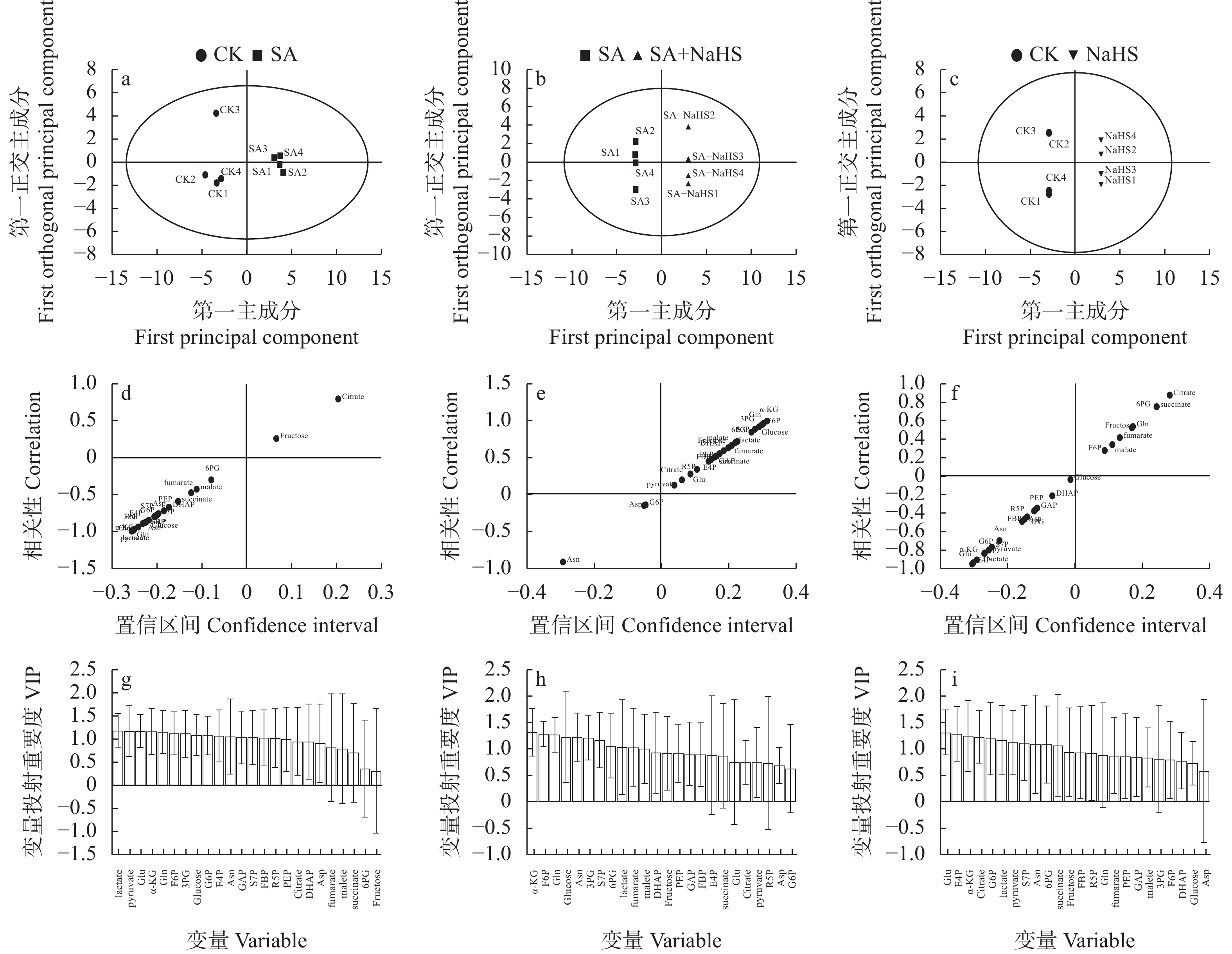
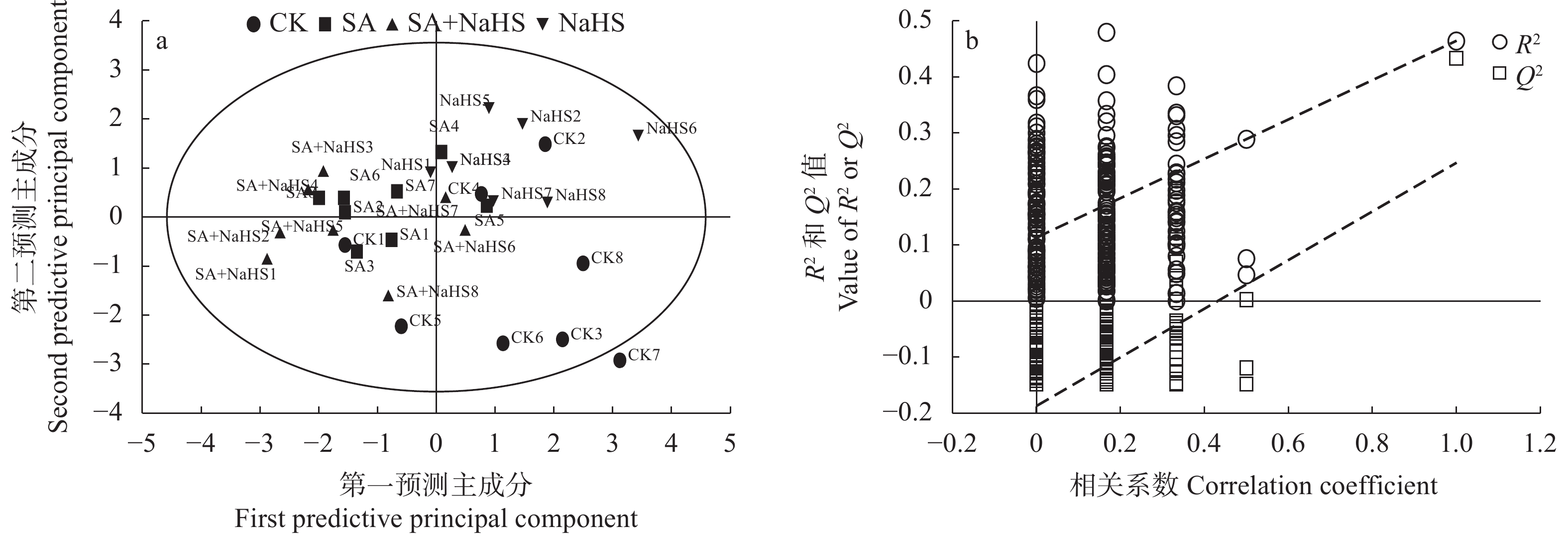
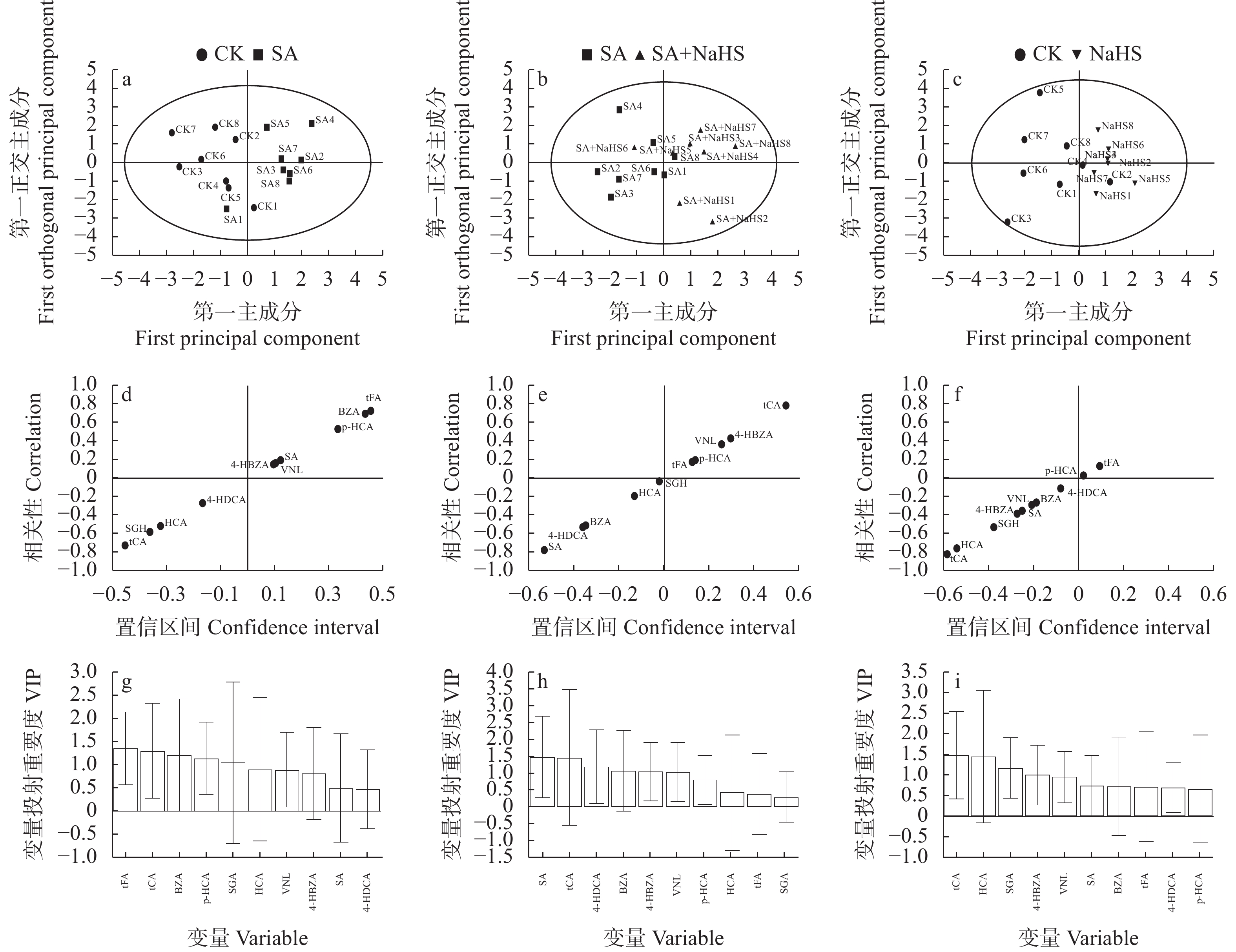
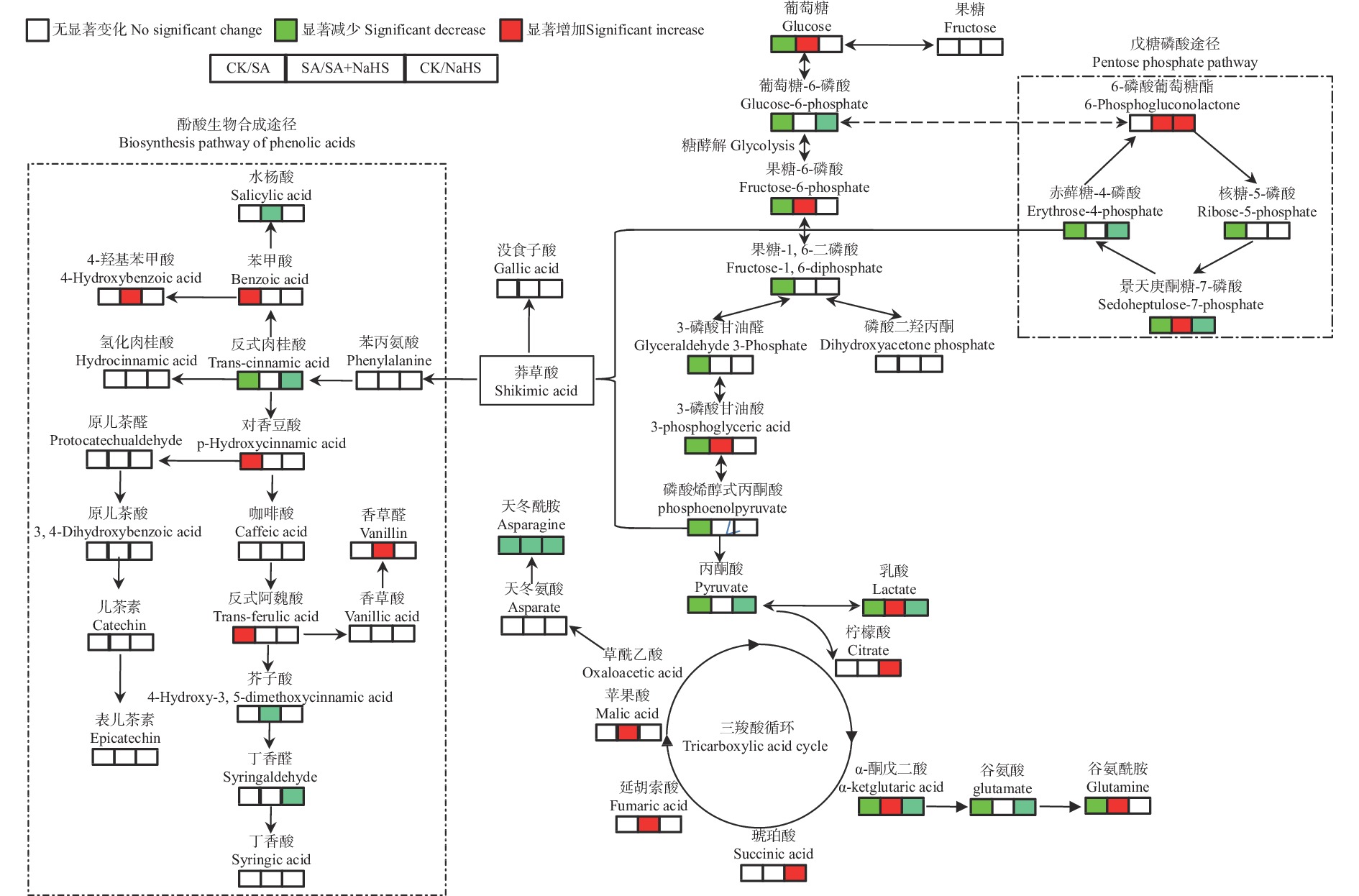
为明确硫化氢信号对盐碱胁迫下植物代谢组的调控作用, 揭示其增强植物耐盐碱性的机理, 以裸燕麦(Avena nuda L.)为材料进行盆栽土培试验, 设置不添加盐碱和添加3.00 g·kg−1盐碱(摩尔比NaCl∶Na2SO4∶Na2CO3∶NaHCO3=12∶8∶1∶9)与裸燕麦抽穗期叶面喷施蒸馏水和50 µmol·L−1硫化氢供体硫氢化钠溶液交叉共4组处理, 利用超高效液相色谱-串联质谱技术结合正交偏最小二乘判别分析方法, 研究外源硫化氢对盐碱胁迫下裸燕麦叶片糖分解代谢产物水平、氧化还原平衡、酚酸含量和产量性状的影响。结果表明: 1)非盐碱条件下, 喷施硫氢化钠对裸燕麦叶片还原型谷胱甘肽/氧化型谷胱甘肽、还原型辅酶Ⅱ/氧化型辅酶Ⅱ、腺苷三磷酸含量和产量性状的影响微弱, 但显著上调柠檬酸、琥珀酸和6-磷酸葡萄糖酯含量, 显著下调葡萄糖-6-磷酸、丙酮酸、乳酸、α-酮戊二酸、谷氨酸、天冬酰胺、赤藓糖-4-磷酸、景天庚酮糖-7-磷酸含量; 盐碱胁迫导致裸燕麦叶片葡萄糖、葡萄糖-6-磷酸、果糖-6-磷酸、果糖-1,6-二磷酸、3-磷酸甘油醛、3-磷酸甘油酸、丙酮酸、乳酸、α-酮戊二酸、谷氨酸、谷氨酰胺、天冬酰胺、赤藓糖-4-磷酸、景天庚酮糖-7-磷酸、核糖-5-磷酸等糖酵解、三羧酸循环和戊糖磷酸途径中间代谢物及还原型谷胱甘肽、氧化型谷胱甘肽、还原型辅酶Ⅱ、氧化型辅酶Ⅱ含量显著降低, 而还原型谷胱甘肽/氧化型谷胱甘肽显著提高; 喷施硫氢化钠显著提高盐碱胁迫下裸燕麦叶片葡萄糖、果糖-6-磷酸、3-磷酸甘油酸、乳酸、α-酮戊二酸、延胡索酸、苹果酸、谷氨酰胺、6-磷酸葡萄糖酯、景天庚酮糖-7-磷酸含量, 显著降低天冬酰胺含量。2)喷施硫氢化钠显著下调非盐碱条件下裸燕麦叶片反式肉桂酸和丁香醛含量; 盐碱胁迫导致裸燕麦叶片反式肉桂酸含量显著降低, 而苯甲酸、对香豆酸和反式阿魏酸含量显著提高; 喷施硫氢化钠显著提高盐碱胁迫下裸燕麦叶片4-羟基苯甲酸和香草醛含量, 显著降低水杨酸、芥子酸含量。3)喷施硫氢化钠对盐碱胁迫造成的裸燕麦穗粒数量和籽粒产量下降具有显著缓解作用, 但对穗数量、穗铃数量、千粒重量和生物学产量的影响不显著。由此表明, 外源硫化氢参与裸燕麦糖分解代谢和酚酸水平调控, 能够增强裸燕麦耐受盐碱胁迫的能力, 它对糖分解途径有机酸水平的提升作用和酚酸独特的调节效应可能在其增强裸燕麦耐盐碱性中发挥着重要作用。
为筛选耐盐高产小麦品种, 确定较为快捷有效的小麦耐盐鉴定指标, 本研究采用田间试验, 以前期筛选出的5个耐盐差异性显著的春性小麦品种(较耐盐品种: ‘宁麦21’ ‘扬麦20’ ‘扬辐麦4’; 盐敏感品种: ‘扬麦23’ ‘安农1124’)为材料, 在江苏大丰地区设置非盐逆境(S0, 播前土壤含盐量0.770±0.062 g∙kg−1)和盐逆境(S1, 播前土壤含盐量3.294±0.198 g∙kg−1)两个处理, 研究了盐胁迫对不同耐盐性小麦花后生理特性及产量的影响。结果表明, 在盐逆境下, 春小麦叶面积指数、干物质积累量及茎蘖数显著下降; 穗数、每穗粒数与千粒重也显著减少, 其中穗数降幅达60.7%, 为减产的主导因素; 其次是粒重的减少, 最终导致产量显著降低, 仅为非逆境条件下的26.2%。研究还发现, 在盐碱胁迫下, 花后SPAD值与叶绿素荧光Fv/Fm降低, 丙二醛与脯氨酸含量显著增加, 变化幅度在品种间存在差异, 耐盐品种的SPAD值及叶绿素荧光Fv/Fm的降幅与丙二醛的增幅较小, 但脯氨酸含量的增幅较大, 因而减产幅度较小。进一步分析表明, 叶绿素荧光Fv/Fm与穗数、千粒重及产量间呈显著正相关关系, SPAD值与干物质积累量呈显著正相关, 干物质积累量与穗数、千粒重及产量间呈显著正相关关系。这些结果表明盐胁迫通过降低叶绿素含量与叶绿素荧光Fv/Fm抑制小麦的光合作用, 减少光合产物的生成, 造成最终的减产。因此在小麦耐盐性田间鉴定中, 开花期SPAD值与叶绿素荧光Fv/Fm可作为耐盐性鉴定快捷可靠的指标。此外, ‘扬麦20’在盐碱地中减产幅度最小, 综合表现较好, 适合在大丰地区盐碱地种植。
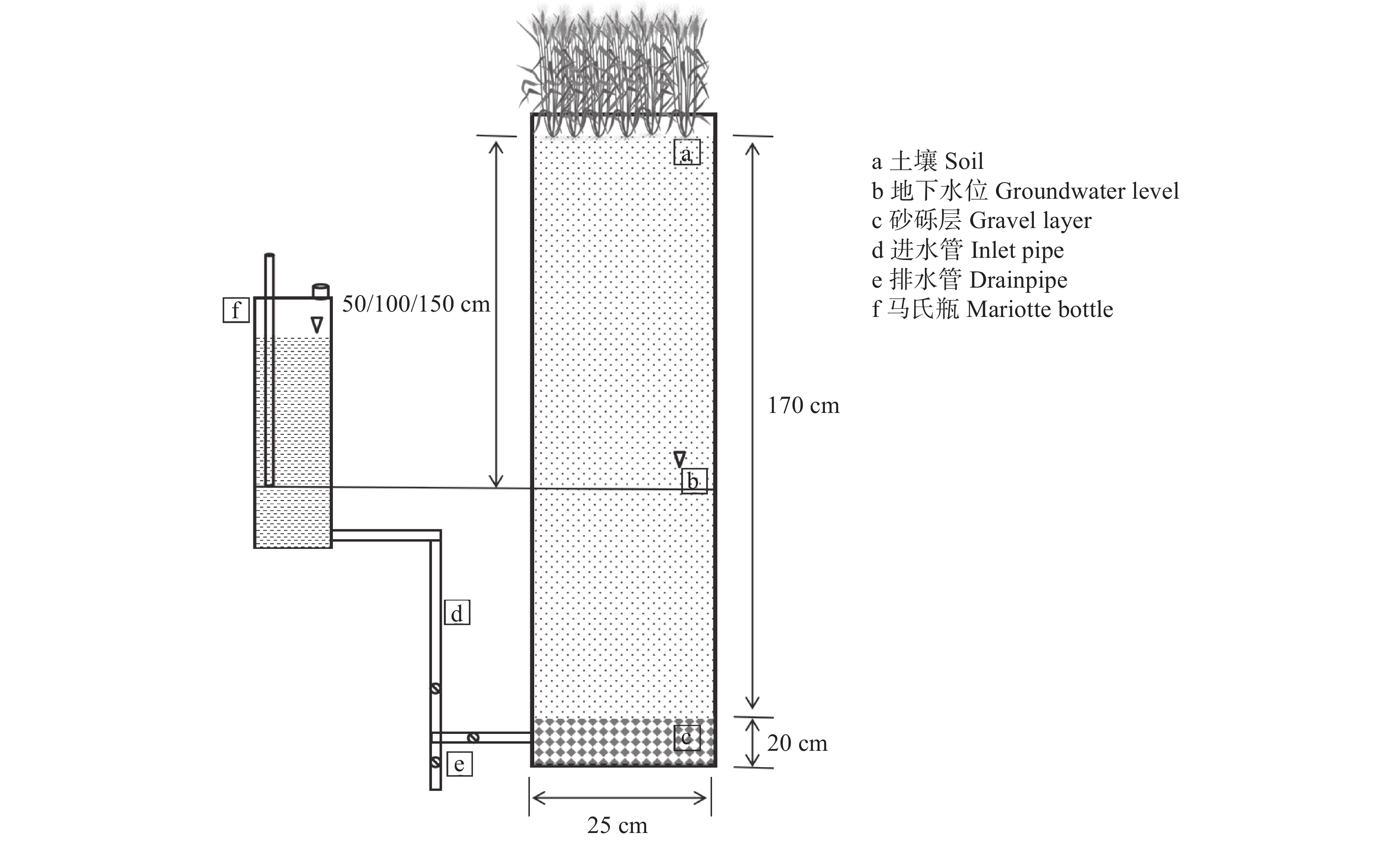
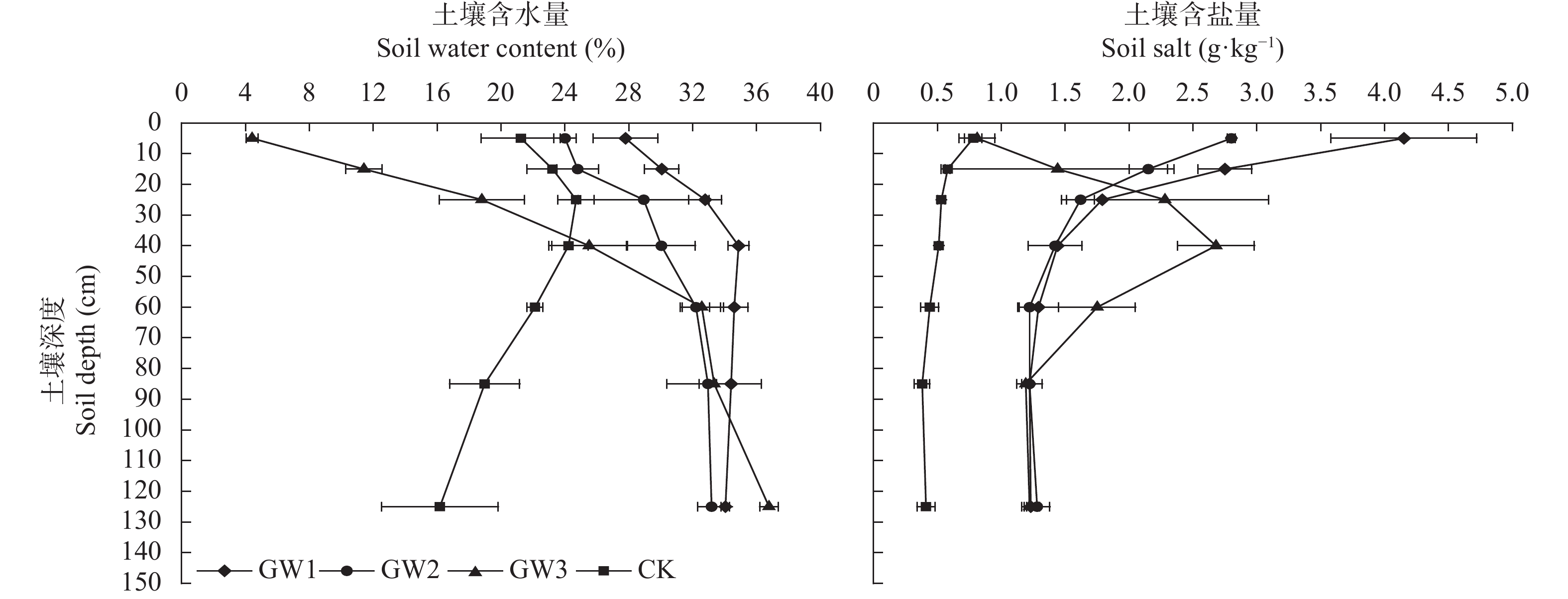
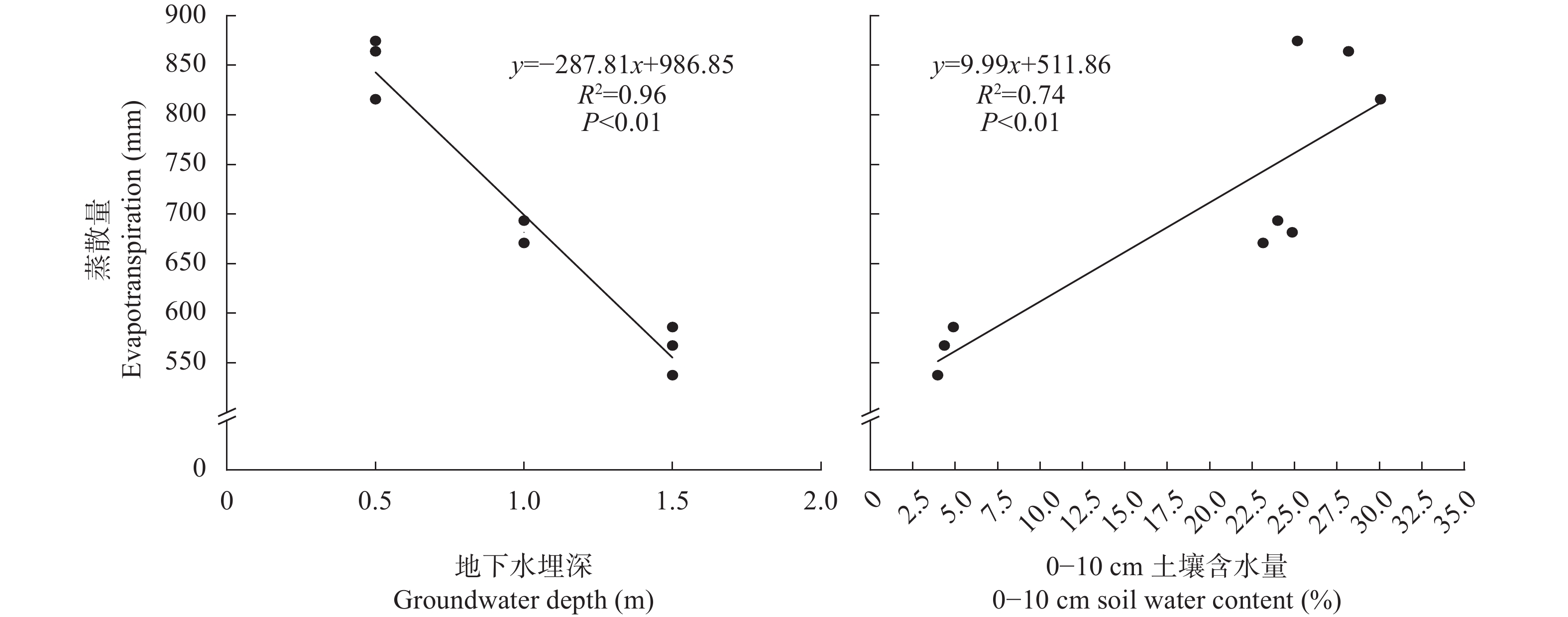
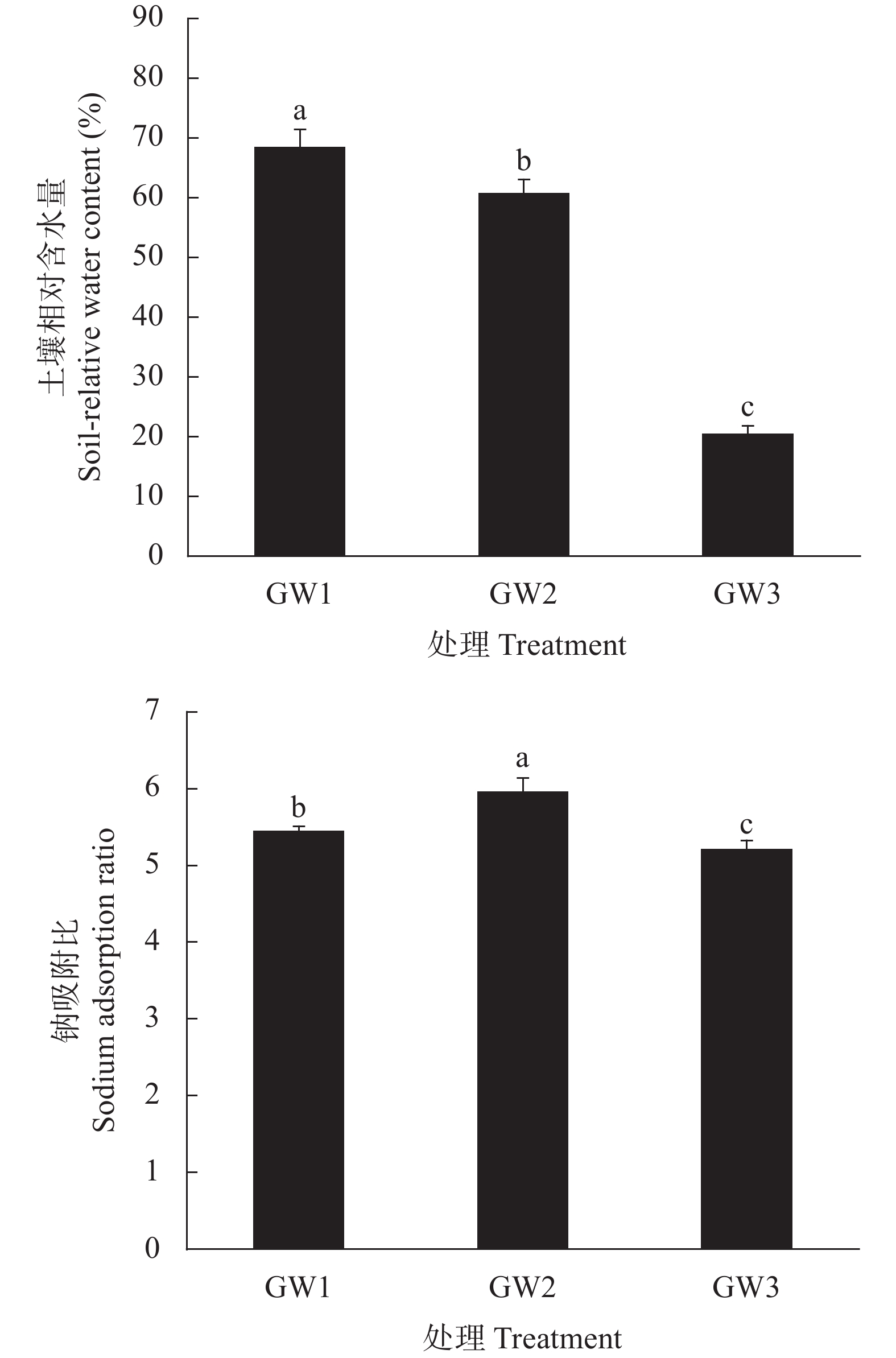
环渤海低平原地区的冬小麦生产面临淡水资源短缺的制约, 高效安全利用较为丰富的浅层微咸水资源对农业可持续发展具有非常重要的意义。本研究于2021—2022年采用土柱模拟方法在中国科学院南皮生态农业试验站开展, 试验设置4个处理: 无地下水埋深淡水灌溉处理(CK), 地下微咸水埋深为0.5 m (GW1)、1.0 m (GW2)及1.5 m (GW3)处理, 每个处理3个重复, 对不同微咸水埋深下冬小麦土壤含水量及含盐量和水分利用特点进行调查。结果表明, 表层土壤(0~10 cm)含水量及含盐量随地下水埋深的增加而逐渐降低。与CK处理相比, GW1处理表层土壤含水量显著增加30.9%, GW3处理显著降低79.3%, 而与GW2处理无显著变化; GW1和GW2处理表层土壤盐含量显著增加3.4 g∙kg−1和2.0 g∙kg−1, 而GW3与CK处理之间差异不显著。GW1和GW2处理盐分主要积累在土壤表层(0~10 cm); GW3处理表层盐分较低, 主要积累在土壤30~50 cm深度。冬小麦蒸散量随地下水埋深的增加而显著降低, 与CK处理相比, GW1和GW2处理下冬小麦蒸散量显著增加50.2%和20.3%, GW3处理无显著差异。冬小麦产量在GW3处理下最高, 较CK处理显著提高38.04%; 同时, 该处理具有最高的生物量水平水分利用效率和产量水平水分利用效率, 显著高于CK处理26.7%和40.1%。上述结果表明, 咸水质量浓度为3 g·L−1, 地下水埋深在0.5~1.5 m的条件下, 1.5 m是冬小麦生长的适宜地下微咸水埋深上限, 此时, 表层含盐量和作物蒸散量最低, 产量和水分利用效率最优。
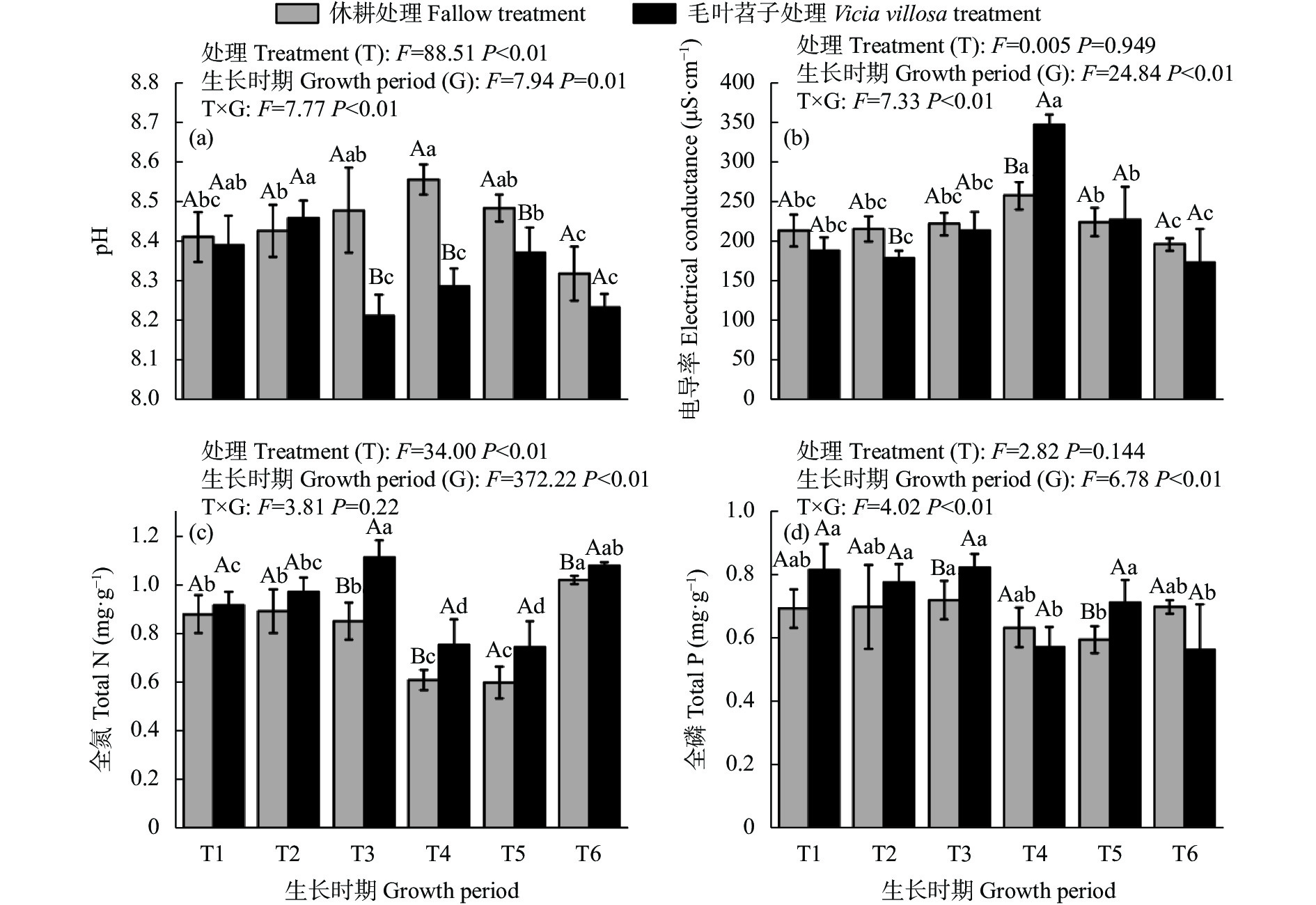
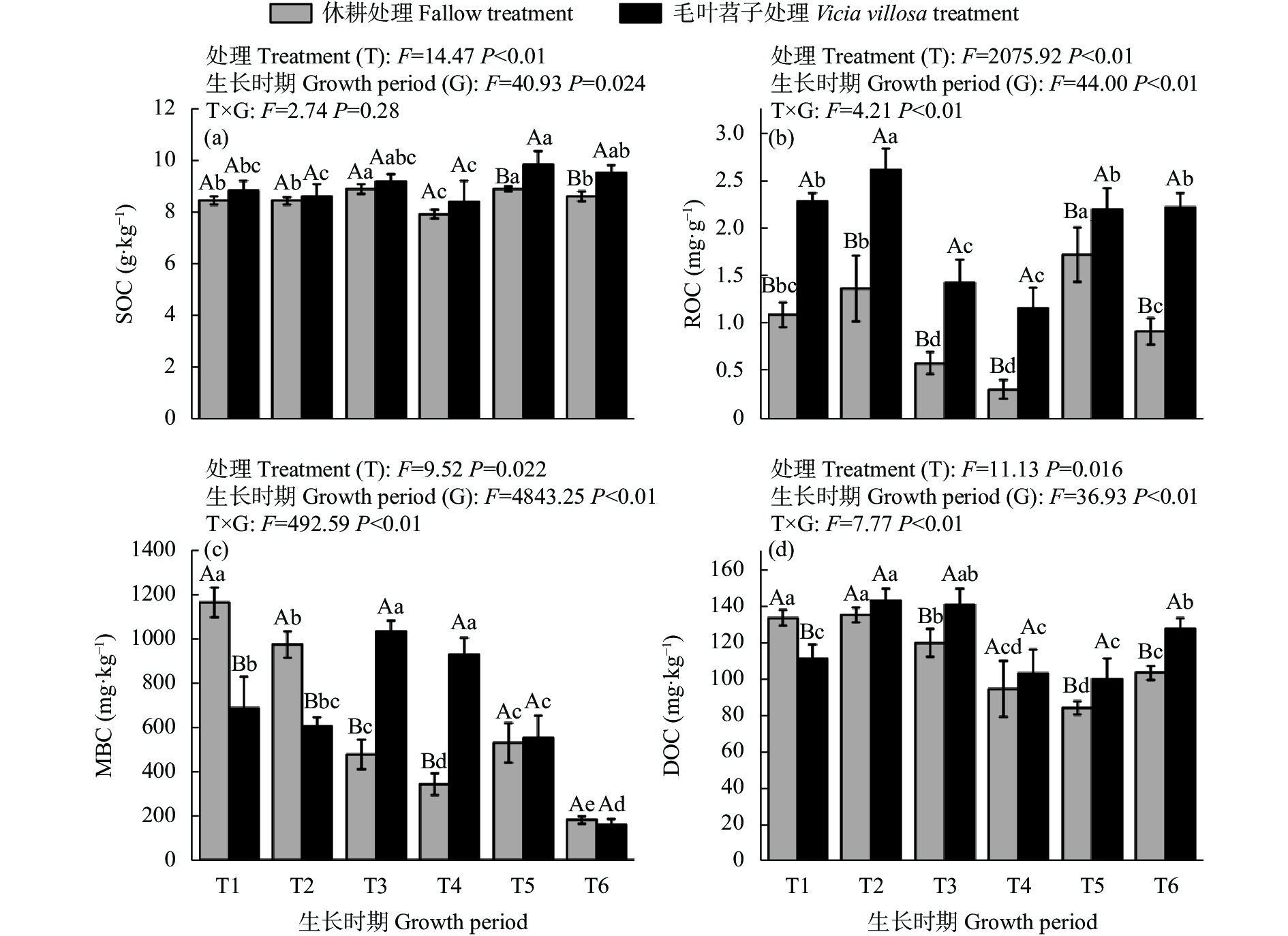
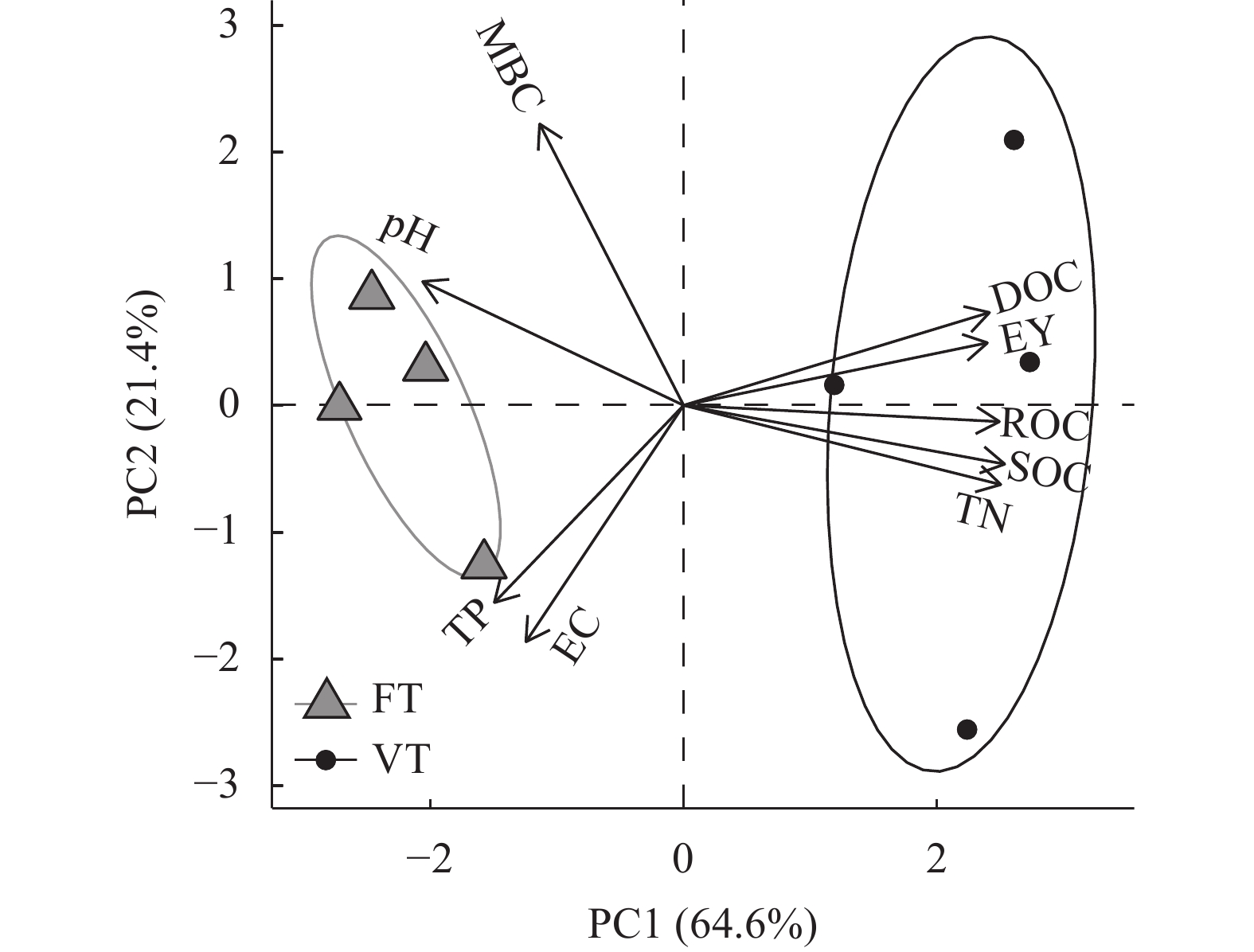
黄河三角洲地区土壤盐渍化严重, 加之冬春季节降雨量少, 淡水资源匮乏, 耕地冬春休耕现象普遍。于2020年9月—2021年10月, 以冬春休耕为对照, 研究种植翻压毛叶苕子对土壤理化性质、活性有机碳组分动态变化和后茬作物玉米产量的影响, 以期为覆盖植物在黄河三角洲地区盐碱地产能提升方面的应用提供参考。结果表明, 在1年试验期内, 与冬春休耕相比, 种植毛叶苕子可以降低土壤EC, 提高易氧化有机碳(ROC)含量, 翻压后则显著降低pH, 并提高土壤养分和活性有机碳含量。与休耕相比, 试验期内毛叶苕子处理平均pH降低0.12, 平均土壤总氮(TN)、总磷(TP)、有机碳(SOC)、ROC、可溶性有机碳(DOC)含量和ROC/SOC分别提高15.1%、5.5%、6.3%、99.1%、8.2%和89.9%, 平均EC则基本持平。毛叶苕子处理的后茬玉米籽粒产量提高15.9%, 增产效果显著。主成分分析结果表明, 玉米产量与土壤TN、SOC、DOC、ROC呈正相关, 与pH、EC呈负相关。毛叶苕子翻压后, 土壤有机碳各组分含量与TN和pH的相关关系增强, 与TN呈显著正相关, 与pH呈显著负相关。种植翻压毛叶苕子后土壤TN升高和土壤pH降低, 提升了土壤有机碳和活性有机碳含量, 综合作用使后茬玉米产量提高。在黄河三角洲地区, 相对于冬春休耕, 冬春季种植毛叶苕子对土壤改良和后茬作物产量提升优势明显, 可考虑作为盐碱地综合利用的优选模式。

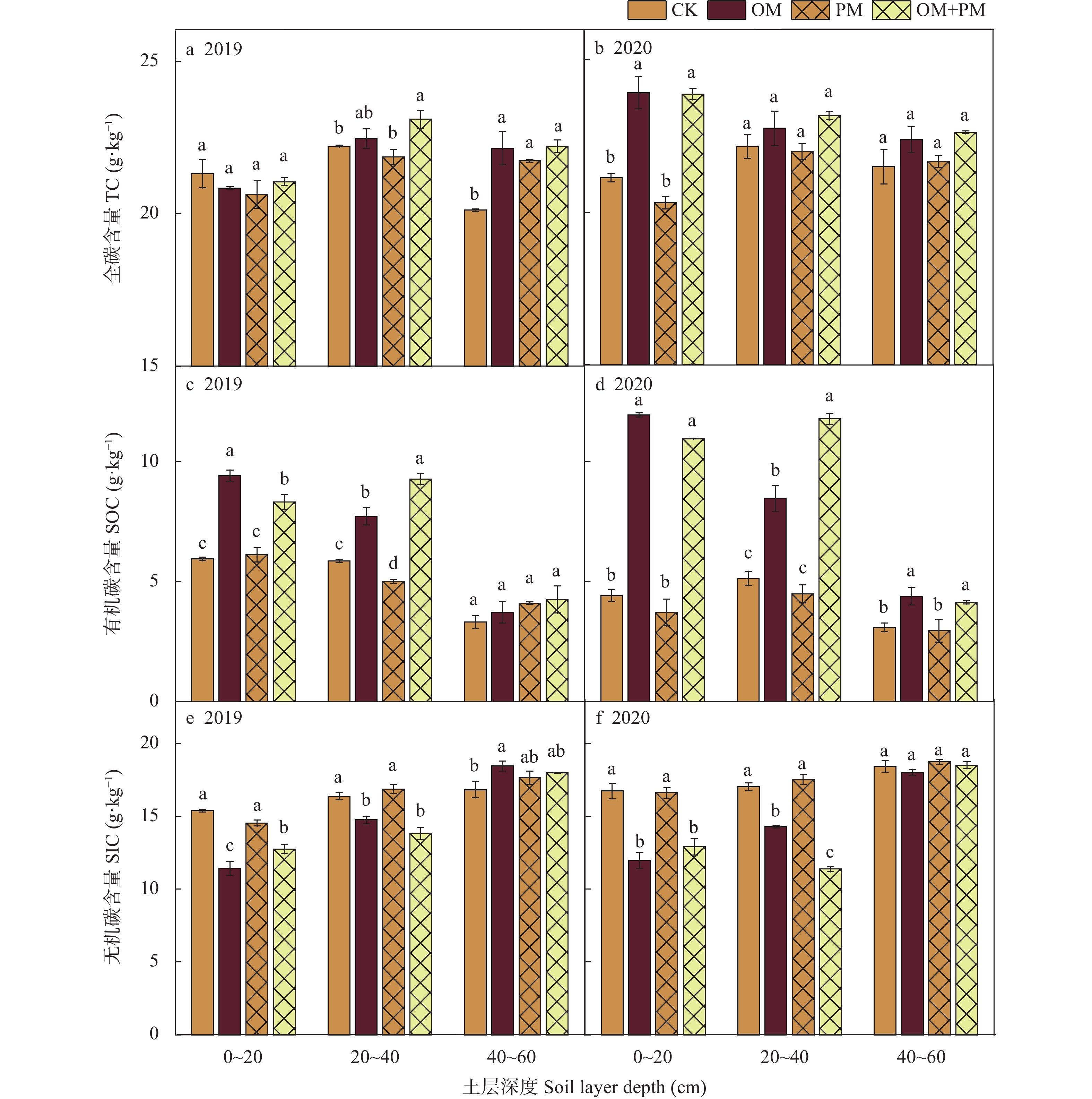
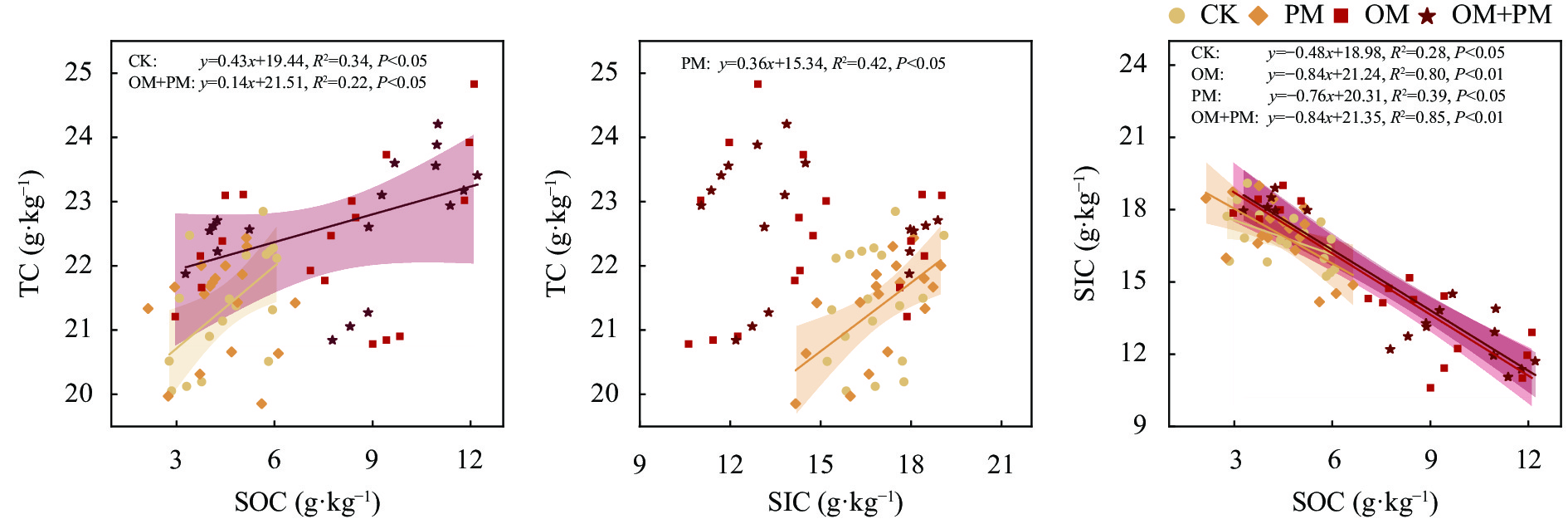
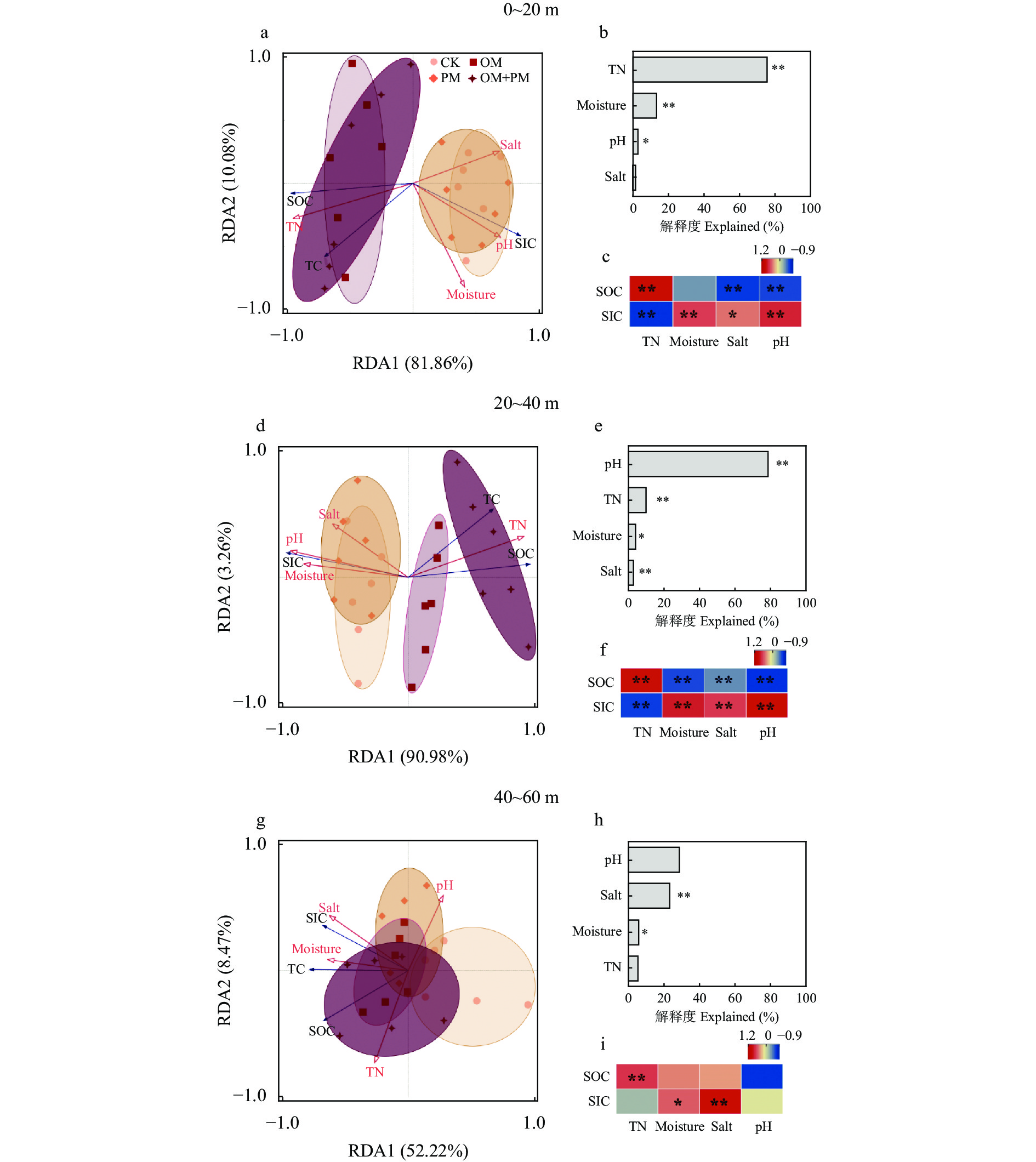
土壤有机碳(SOC)和无机碳(SIC)是参与全球碳循环的重要碳库。亚表层(10~30 cm)培肥结合地膜覆盖措施是干旱区优化盐碱土壤物理结构和调控土壤水盐环境的有效措施, 然而关于其如何调控0~60 cm土体SOC、SIC分布及其与土壤相关理化性状的关系尚不明确。本研究基于内蒙古河套灌区盐碱土壤6年的田间微区试验, 设置常规对照(CK)、亚表层(10~30 cm)有机培肥(OM)、地膜覆盖(PM)、亚表层有机培肥+地膜覆盖(OM+PM) 4个处理, 测定了2019—2020年0~60 cm剖面SOC、SIC、全碳(TC)含量以及土壤理化指标(土壤水分、盐分、pH和全氮), 分析了TC、SOC、SIC变化特征及其影响因素。结果表明: OM和OM+PM处理较CK和PM处理显著增加0~40 cm土层SOC含量31.9%~195.6% (P<0.05), 显著增加40~60 cm SOC含量33.7%~49.4% (P<0.05, 仅2020年), 但显著降低0~40 cm SIC含量9.9%~35.0% (P<0.05)。基于SOC和SIC的变化, OM+PM较CK处理显著增加2019年20~60 cm TC含量10.4%~39.4% (P<0.05), 并显著增加2020年0~20 cm TC含量13.0% (P<0.05)。回归分析结果进一步说明, 覆膜条件下补充亚表层培肥, 使总碳库变化的主导因素由SIC转变为SOC。冗余分析结果表明土壤理化性质是影响土壤碳库的主要因素(解释度为60.7%~91.9%), 其中全氮和pH是0~40 cm土壤碳库的主要影响因子, 而40~60 cm土壤碳库主要受盐分和pH影响。相关性分析结果表明SOC与SIC表现为完全相反的变化规律, 其中SOC与全氮极显著正相关, 与盐分和pH呈极显著负相关(P<0.01); SIC与全氮呈极显著负相关, 与pH呈极显著正相关(P<0.01)。因此, 亚表层培肥结合地膜覆盖可以通过增加SOC来弥补SIC的损失进而实现碳积累, 是该区域盐碱地增加固碳潜力的有效措施。
我国约有9913万hm2盐碱地, 其中现代盐渍土约为3693万hm2, 作为重要的后备耕地资源, 盐碱地的综合利用对保障国家粮食安全具有重要的战略意义。经过几十年的研究和实践, 我国在盐碱地治理方面已经形成了完善的理论和技术体系, 取得巨大的成就, 其中淡水洗盐、灌排结合的水利工程措施在盐碱地治理中发挥了关键作用。但是, 受水资源短缺和气候条件制约, 我国大部分的盐碱地尚未开发利用。近年来, 随着社会经济发展和科学技术进步, 人们逐渐认识到盐碱地作为自然生态系统的组成部分, 其丰富的盐碱土、咸水和耐盐生物等盐碱资源在多元化食物生产、生态保护、人民生活中具有重要作用, 迫切需要加强盐碱资源高效利用的理论与技术研究, 发展盐碱地资源高效利用产业, 推动盐碱区农业高质量发展。本文分析了目前盐碱地治理现状及其面临的问题, 结合盐碱地资源高效利用研究进展, 重点对盐碱区盐生植物利用、咸水资源的高效利用、肥沃耕层构建高效改土、高品质农产品生产、设施种养空间利用和草牧业发展等六个方面进行了探讨, 以期为盐碱地资源高效利用提供参考。
苏打盐碱土作为我国后备土地资源, 具有极大利用潜力, 若加以合理的开发利用, 将会产生巨大收益。在苏打盐碱土改良中, 化学方法是一种重要的手段, 主要包括含钙制剂、有机类改良剂和酸性物质, 其中含钙制剂和有机类改良剂主要通过添加外源钙置换土壤胶体吸附的交换性钠和改善土壤结构促进水分淋洗来改良苏打盐碱土。苏打盐碱土自身富含碳酸钙, 以酸性物质作为化学改良剂可发生水解产生氢离子, 降低盐碱土pH, 有利于溶解土壤中的碳酸钙, 为代换土壤中的交换性钠提供钙源, 从而减少外源钙质材料添加, 降低改良成本。本文通过综合分析国内外酸性物质对苏打盐碱土物理性质、盐碱性质、养分利用和作物产量等方面的研究, 对酸性物质应用于苏打盐碱土改良的机理与实践进行总结, 并展望其未来发展趋势, 以期为苏打盐碱土治理与农业利用提供参考。
水资源是基础性自然资源和重要的战略资源。盐碱地多分布于干旱半干旱地区, 淡水资源短缺是盐碱区农业可持续发展的主要限制因素。同时盐碱区较为丰富的咸水微咸水资源、土地资源和光热资源等为盐碱区农业可持续发展提供了可能。本文针对咸水灌溉影响耕地质量、作物生长、产量和品质等问题, 综述了基于水质的咸水分类、咸水灌溉制度与灌溉方式和地下水埋深等影响咸水在农业生产中安全利用的因素, 阐述了不同矿化度咸水灌溉, 土壤水力特性、理化性质、温室气体排放等土壤质量变化情况和对作物生长发育、产量和品质的影响, 明确了有机物料、咸水灌溉制度、覆盖和耕作等农艺措施、水肥盐多因素调控和耐盐作物适盐种植等农业措施的作用。咸水灌溉下土壤质量呈下降趋势, 有机物料的施用、秸秆还田和合理的耕作等调控措施通过影响土壤质量保证咸水的安全利用。与旱作相比, 咸水灌溉可以起到明显的增产作用, 在合理的咸水范围内还能提升品质。在新形势下, 未来将面向国家粮食安全重大需求, 以协同提升土壤质量、作物产量和品质为多目标, 系统开展咸水非充分灌溉、水肥盐综合调控、咸水灌溉对土壤质量和作物咸水精准灌溉机理过程研究、技术研发和模式示范工作, 为缺水盐渍区农业可持续发展提供理论依据和技术支撑。
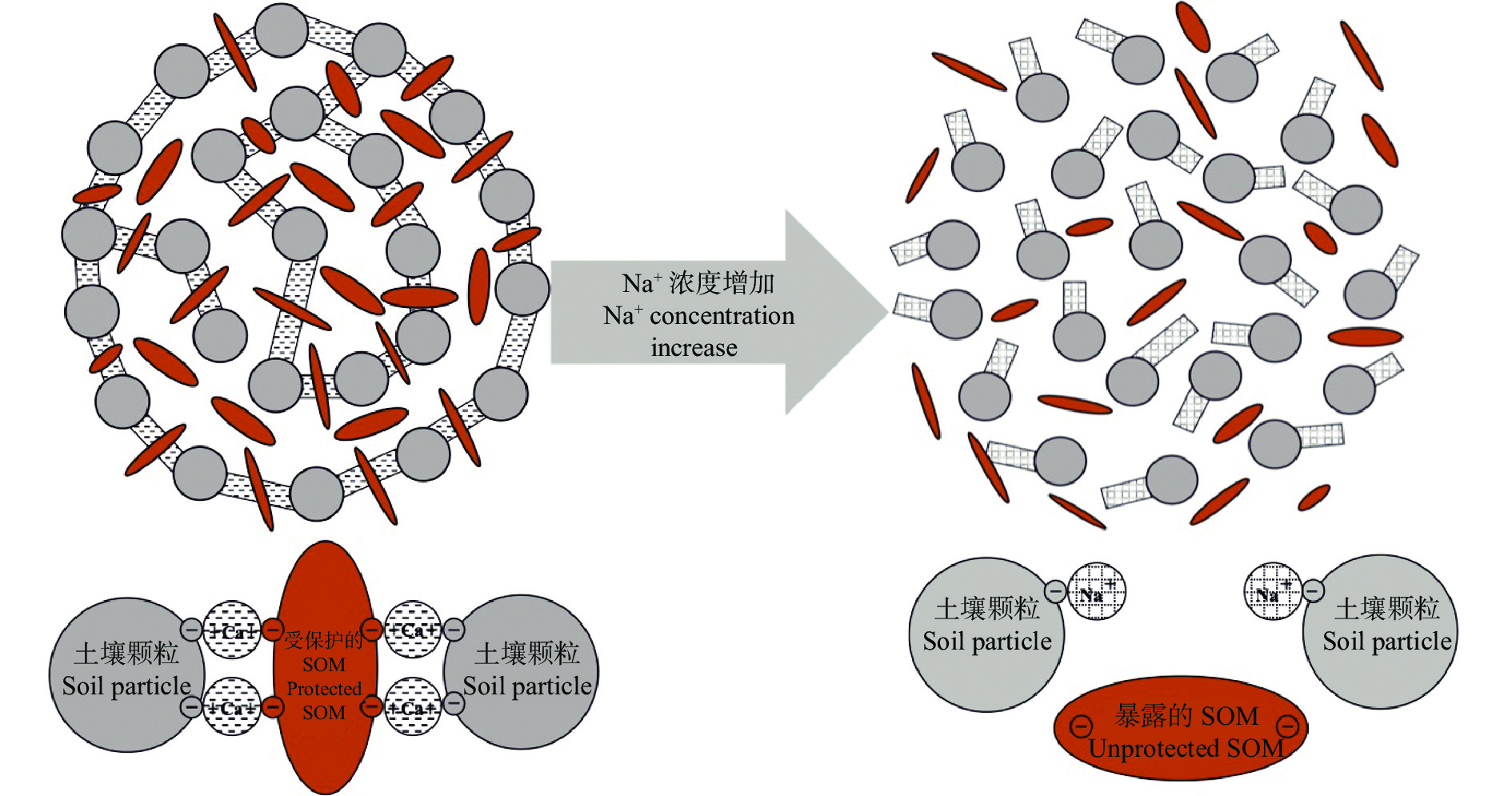
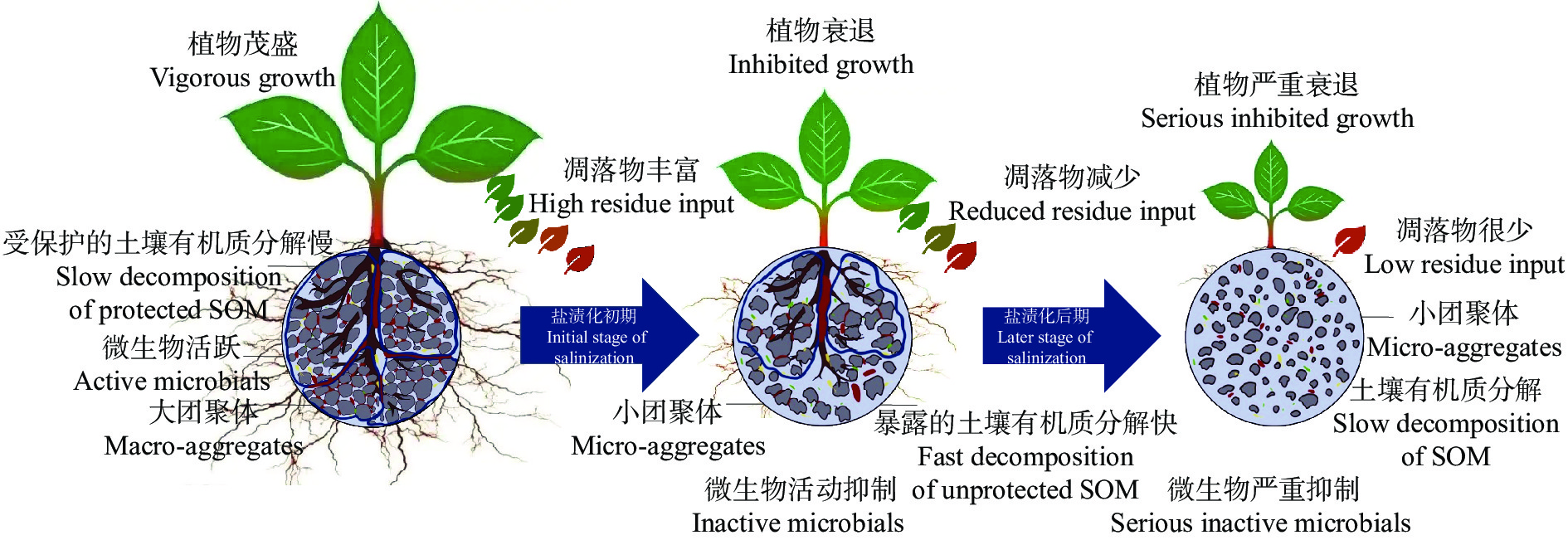
土壤有机质是耕地质量的核心, 不仅促进土壤团聚体形成, 也为植物和微生物提供养分。土壤有机质的形成和分解过程都离不开微生物的参与, 而土壤团聚体不仅为微生物提供了栖息环境, 也对有机质进行物理保护。在高盐分含量的土壤中, 有机质的积累和分解过程变得更加复杂, 因此本文总结了土壤盐渍化及其危害, 分析了土壤盐分对土壤团聚结构和微生物特征的影响、盐碱土壤有机质特征及积累规律, 进而综述了土壤盐分对土壤有机质影响规律的研究进展, 旨在揭示盐碱土壤碳封存的潜在机理。以往研究表明盐渍化土壤有机质含量低、团聚结构差、微生物活性低, 这些都与土壤盐分含量高和外源有机物质输入量低有关。恶劣的土壤结构导致盐渍化土壤有机质暴露而较易分解, 低量的外源有机物质输入导致盐渍化土壤有机质较难积累。可见, 盐渍化土壤是潜在的碳库, 适宜的措施可以显著提高盐渍化土壤有机质含量。在此基础上, 提出未来盐渍化土壤有机质积累的研究方向: 1)不同盐分环境下土壤团聚结构和土壤微生物在有机质分解过程中的响应规律; 2)外源有机物料添加下土壤团聚结构和土壤微生物在有机质积累过程中的响应规律; 3)土壤有机质提升后盐碱地生产力特征。以上研究不仅可以阐明盐碱土壤有机质的周转机理, 为盐碱地“固碳封存”提供理论依据, 还可以有针对性地提供盐碱耕地质量提升措施, 推进盐碱地绿色可持续发展。
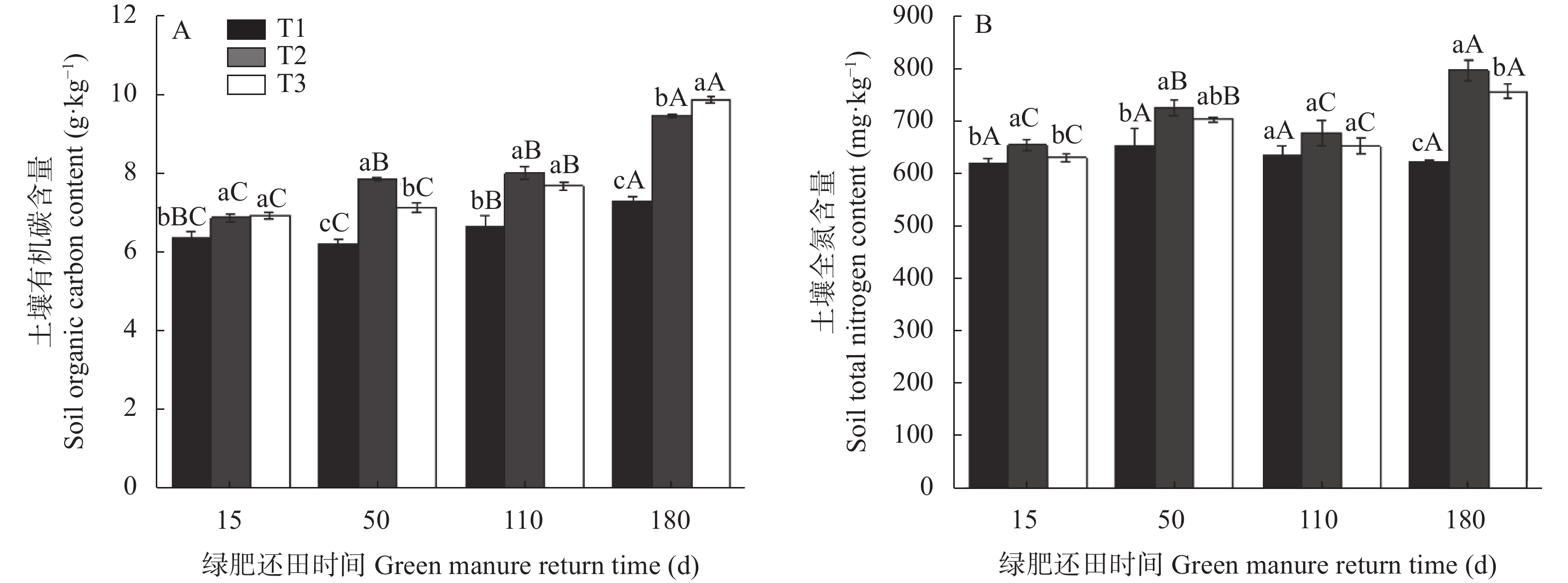
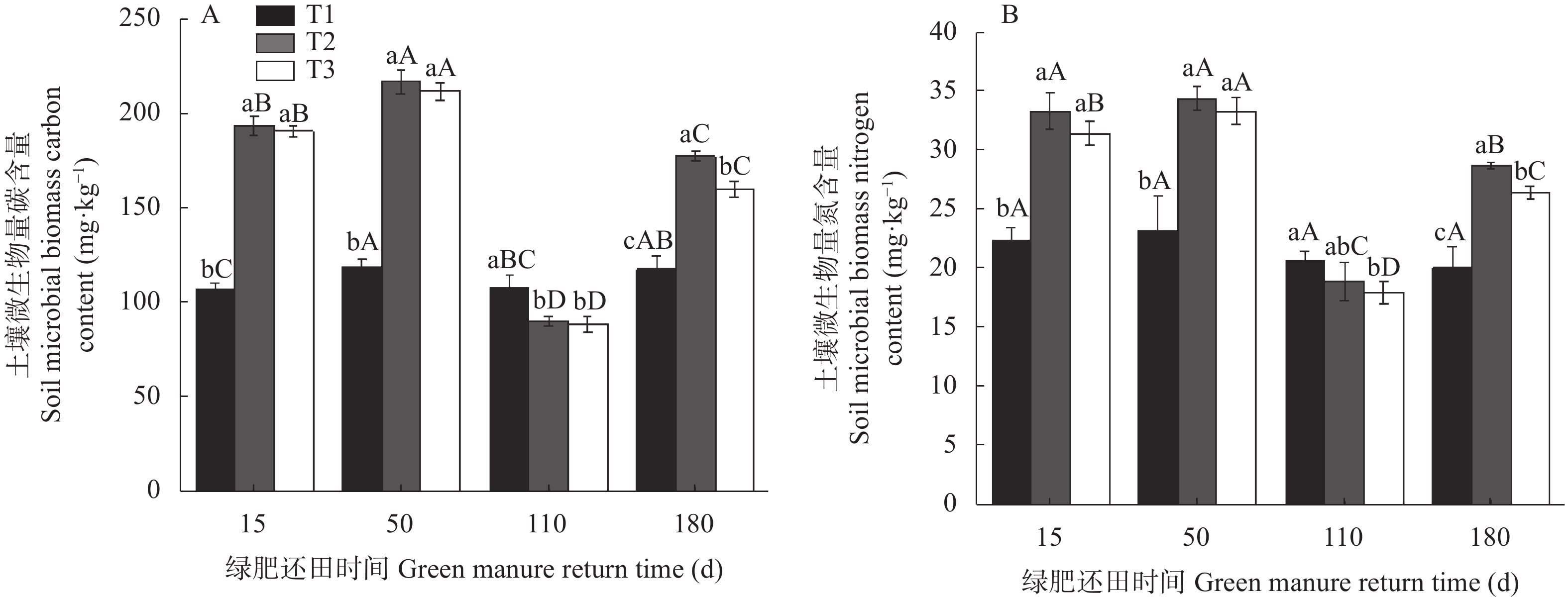
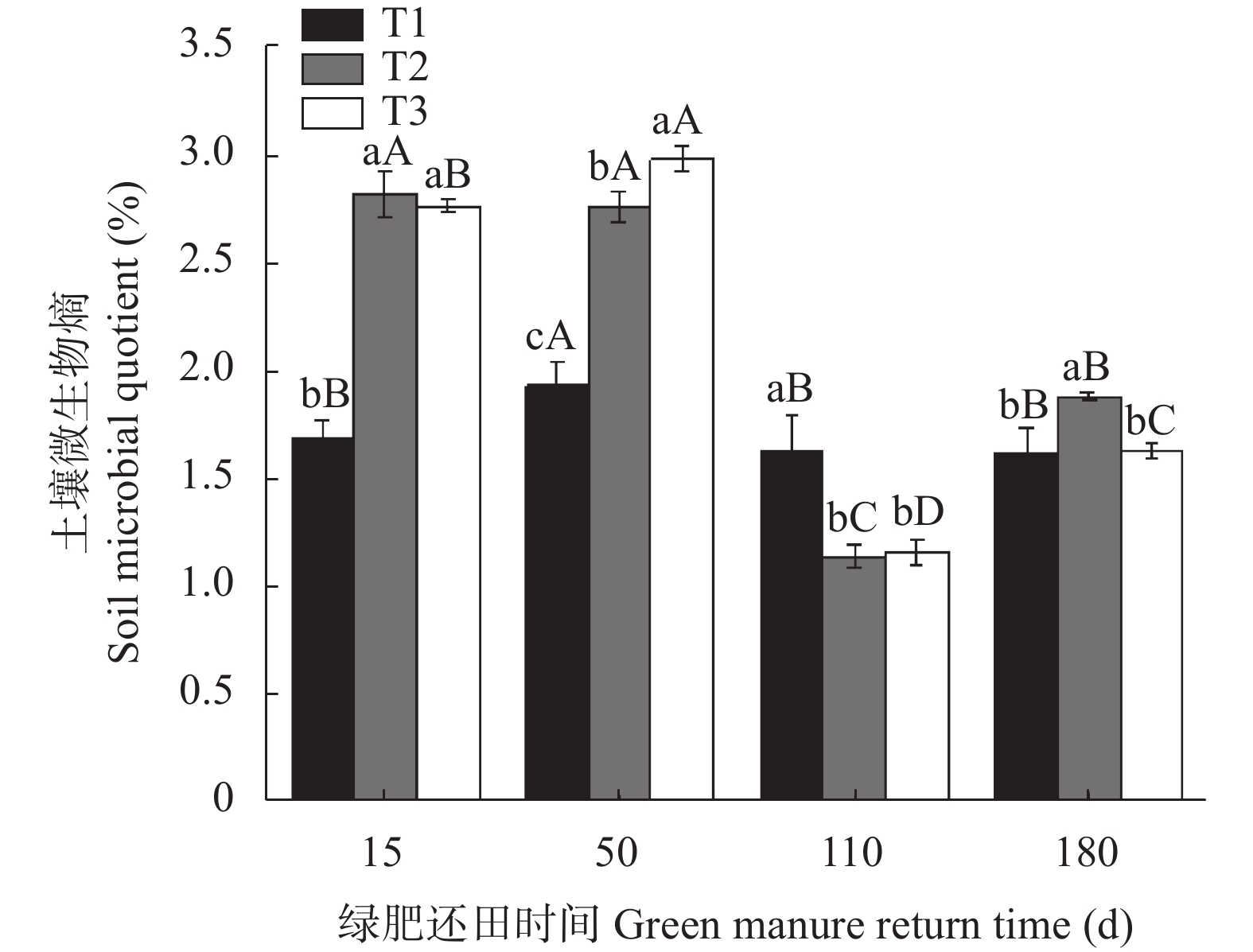
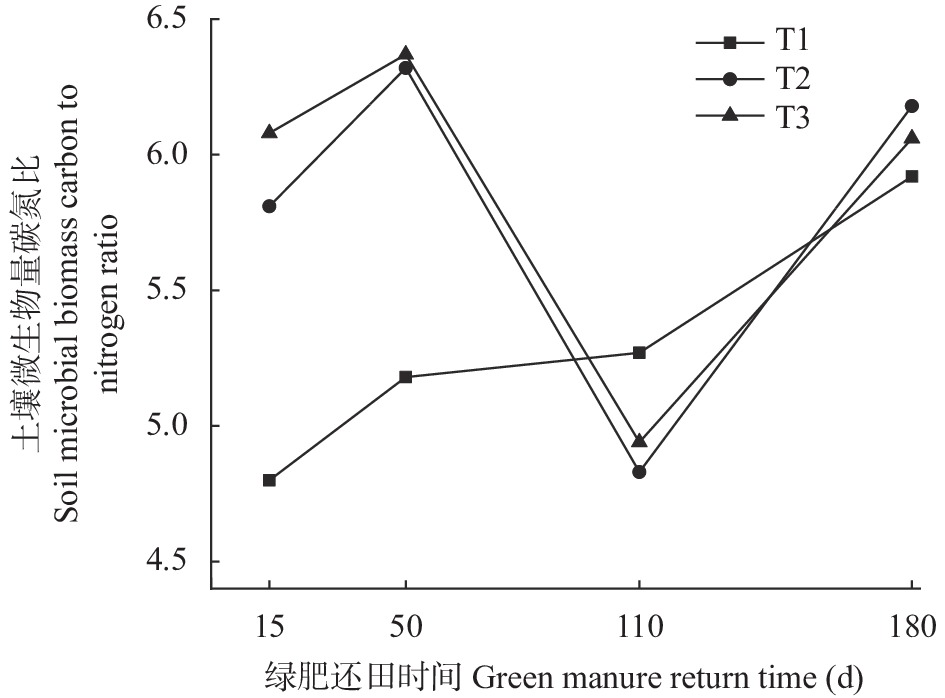
为探讨绿肥还田对盐碱地棉田土壤碳氮和微生物量碳氮的影响, 于2018—2019年选取黑麦草‘冬牧70’和大麦‘驻大麦4号’ 2种耐低温耐盐碱的禾本科绿肥进行原位还田试验, 设置冬闲农田-棉花(T1)、黑麦草-棉花(T2)和大麦-棉花(T3) 3个处理, 测定不同处理和绿肥还田后不同时期(15 d、50 d、110 d和180 d)棉田土壤有机碳(SOC)、土壤全氮(TN)、土壤微生物量碳(SMBC)和微生物量氮(SMBN)的含量, 并计算土壤微生物熵(SMQ)和土壤微生物量碳氮比(SMBC/SMBN)值。结果表明, T2和T3均能显著增加SOC、TN含量, 并在180 d时达最大值9.50 g∙kg−1、798.84 mg∙kg−1和9.91g∙kg−1、759.34 mg∙kg−1, 分别显著高于T1处理。T2和T3的SMBC、SMBN含量在整个还田期的变化动态基本一致, 呈前期稳定增长且显著高于T1, 后期有所降低且在110 d时略低于T1的变化动态; 并且均在50 d时达最大值, 此时较T1分别高出81.46%、47.76%和77.33%、43.13%; 同时还田后T2处理不同时期的SMBC和SMBN含量均高于T3。SMQ不同处理的变化趋势与SMBC一致, 2种绿肥处理除110 d外均高于T1处理, T2在15 d时达到最大值2.82%, 而T3在50 d时达到最大值2.98%。各处理SMBC/SMBN值均在4~7之间变化, 由此可判断绿肥还田后土壤中微生物群落以细菌为主; 同T1相比, 除110 d外T2和T3均表现出较高的SMBC/SMBN值。综上所述, 在盐碱地冬闲农田种植绿肥并还田可以显著提高棉田土壤碳氮和土壤微生物量碳氮含量, 改善土壤微生物群落组成和提高土壤微生物固碳效应, 为后茬作物生长提供养分。研究结果对盐碱地冬闲田的合理利用具有指导意义。
作物病虫害严重影响粮食产量和质量, 对农业生产造成巨大损失。开展作物病虫害生境适宜性评价能够对适合病虫害繁殖和流行的环境区域进行有效表征, 为病虫害预测提供重要信息。由于作物病虫害发生和流行受多种生境因素影响, 同时这些生境因素时空异质性高, 难以通过传统气象站点数据、人为调查等方式进行有效表征, 为病虫害生境评价带来较大的挑战。遥感技术的发展和成熟为病虫害生境信息表征带来重要机遇。多源遥感信息在时空异质信息表征方面具有天然优势, 同时能与传统气象站点数据形成信息互补, 为病虫害生境适宜性评价提供全面、丰富的信息, 支持生境适宜性评价模型的构建。本文对多源遥感信息在作物病虫害生境适宜性评价方面的研究进展进行综述, 重点分析多源遥感数据在寄主作物分布及生长状态、环境气象条件和景观等病虫害生境因子表征方面的潜力, 以及大范围生境适宜性评价涉及的统计模型、机器学习模型和生态位模型等建模方法。在此基础上, 提出基于多源遥感信息的作物病虫害生境评价模型构建的框架, 并对技术的发展趋势进行探讨, 为更加精准、科学的区域尺度病虫害防控管理提供技术支撑, 为病虫害统防统治和绿色防控提供科学指导。
再生稻模式在湖北省粮食生产调结构转方式中具有重要作用, 研究氮肥与有机肥配施对再生稻稻田土壤肥力性状的影响, 可为土壤肥力维持和再生稻高效生产提供科学依据。大田试验于2020—2021年进行, 各试验处理磷(P2O5)、钾(K2O)养分施用量分别为 75 kg?hm?2和150 kg?hm?2, 氮(N)施用量200 kg?hm?2(不包括不施氮处理N0)。按氮肥与有机肥施用情况分为5种基肥处理: 不施氮肥(N0); 基肥氮(N 75 kg?hm?2)全部来自常规尿素(CK); 两种物料配施时, 基肥氮由2种物料各提供一半, 2种物料配施包括缓释尿素与常规尿素(T1)、生物炭与常规尿素(T2)、畜牧粪便与常规尿素(T3)。T2处理区在2021年不再施入生物炭, 施肥与CK处理相同。结果表明: 施入生物炭和畜牧粪便都能降低土壤容重, 以前者效果更佳; 施入生物炭后第1年, 土壤pH、有机碳和全氮明显提高, 第2年的效果与畜牧粪便无明显差异; 在头季稻分蘖期、抽穗期和再生稻抽穗期, 土壤无机氮含量分别以施常规尿素、缓释尿素和畜牧粪便最高; 畜牧粪便和生物炭可提高土壤微生物量碳和微生物量氮含量, 其中在头季稻拔节期前生物炭的施用效果较好, 拔节期后以畜牧粪便的施用效果较好。此外, 在畜牧粪便处理下, β-葡萄糖苷酶和脲酶活性较高。比较而言, 畜牧粪便在降低土壤容重、提高有机碳和全氮的效果上次于生物炭, 在提高无机氮、微生物生物量和土壤酶活性上效果优于生物炭, 因此, 建议基肥采用畜牧粪便与化肥配施, 由畜牧粪便取代其中50%的化肥氮。
在农业景观中非耕作生境通常会为地表节肢动物提供适宜的生存场所, 步甲和蜘蛛群落作为天敌类地表节肢动物的重要组成部分, 非耕作生境类型对其多样性和空间分布的作用有所差异。本文在辽宁省昌图县调查了非耕作生境(即果园、草地、乔木林地和其他林地生境)以及其相邻耕地内部和耕地边缘的步甲和蜘蛛个体数、物种数和多样性, 采用方差分析、聚类分析和冗余分析等方法, 详细讨论了不同生境内步甲和蜘蛛群落分布特征以及非耕作生境类型对邻近耕地边缘及内部步甲和蜘蛛多样性的影响机制。结果表明: 1)不同非耕作生境类型对耕地内部溢出效应明显, 但产生的影响有所差异, 草地对耕地内部步甲多样性影响显著, 其他林地对耕地内部蜘蛛的多样性影响显著。2)不同非耕作生境类型产生的边缘效应与溢出效应规律一致, 均会促进耕地内部步甲和蜘蛛的多样性; 步甲的多样性在邻近草地的耕地边缘显著高于其他边缘类型, 蜘蛛的多样性在邻近其他林地的耕地边缘显著高于其他边缘类型。3)非耕作生境的植被盖度和多样性显著影响步甲和蜘蛛的多样性。总体而言, 非耕作生境对提升耕地内天敌多样性有积极作用, 林地和草地作用效果更为明显, 并且非耕作生境内植被群落结构越复杂, 越有利于维持较高的天敌多样性, 进而提高生物控害功能。因此, 在农业景观中, 增加非耕作生境景观建设,如设置合适农田边界带、种植合适的植被群落等,可以促进农业景观中的生物控制服务, 提升害虫防治功能维持生态系统稳定,对建设高标准农田具有重要意义。
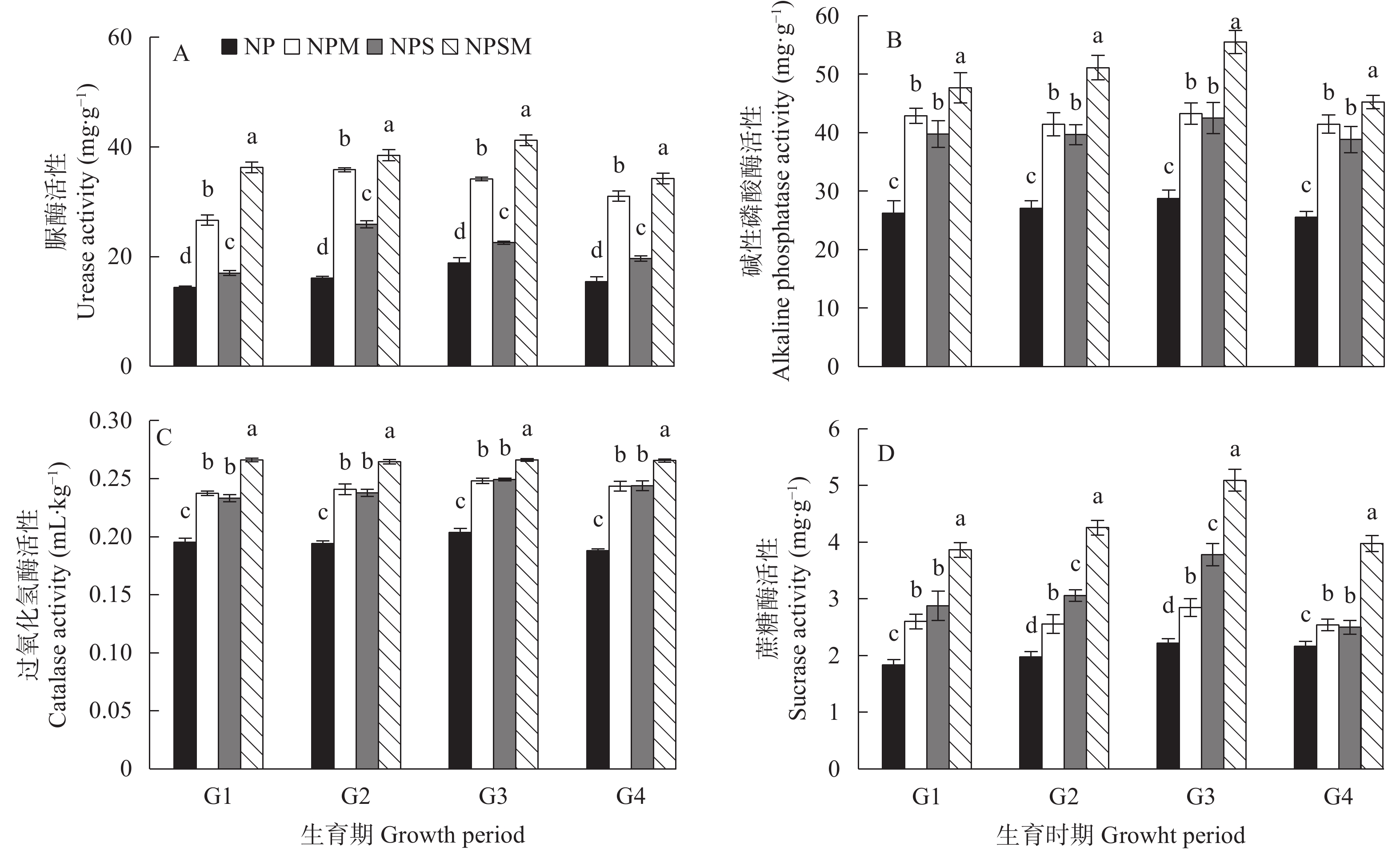
为明确棉花秸秆还田与施用有机肥对土壤的影响, 探究长期棉花秸秆还田和施用有机肥对连作棉田土壤养分含量和酶活性的影响,为棉秆还田和施用有机肥的推广提供依据。 本研究依托山西农业科学院棉花研究所始于2007年牛家凹棉花长期定位试验站, 设置了氮磷化肥+秸秆清茬(NP)、氮磷化肥+秸秆还田(NPS)、氮磷化肥+有机肥(NPM)及氮磷化肥+秸秆还田+有机肥(NPSM)共4个处理, 测定不同处理在棉花不同生育期对土壤养分含量和酶活性的影响, 并对两者之间的关系进行了探究。结果表明: 与处理NP相比, 处理NPS、NPM和NPSM的棉花产量分别显著增加10.23%、11.10%和26.22% (P<0.05)。处理NPSM对土壤养分含量提升幅度最大, 与同期处理NP相比, 全氮、全磷、碱解氮、速效钾和有效磷含量分别显著提高48.76%~60.18%、91.23%~112.18%、140.63%~229.17%、35.86%~60.54%和146.92%~483.34% (P<0.05), 均达到显著性差异水平。同样, 各处理中, 处理NPSM明显提高了土壤的酶活性, 与同期的处理NP相比, 土壤脲酶、磷酸酶、过氧化氢酶和蔗糖酶活性分别显著提高118.50%~151.84%、76.81%~93.11%、30.53%~41.33%和83.41%~129.32% (P<0.05)。相关性分析表明, 土壤养分含量的增加会促进酶活性的提升。因此, 长期秸秆还田配施有机肥后可显著提高连作棉田产量, 增加土壤的养分含量, 提升土壤中酶活性, 从而达到优化土壤结构、提升土壤肥力的目的。
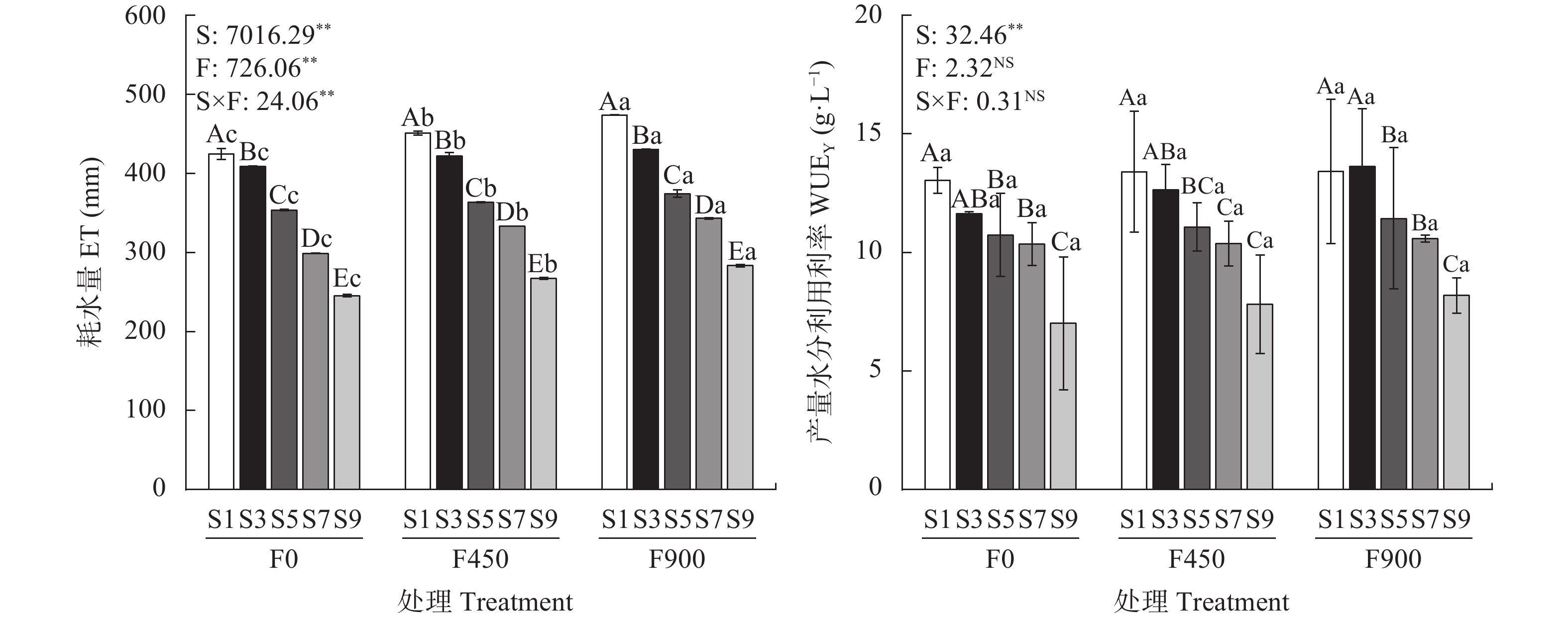
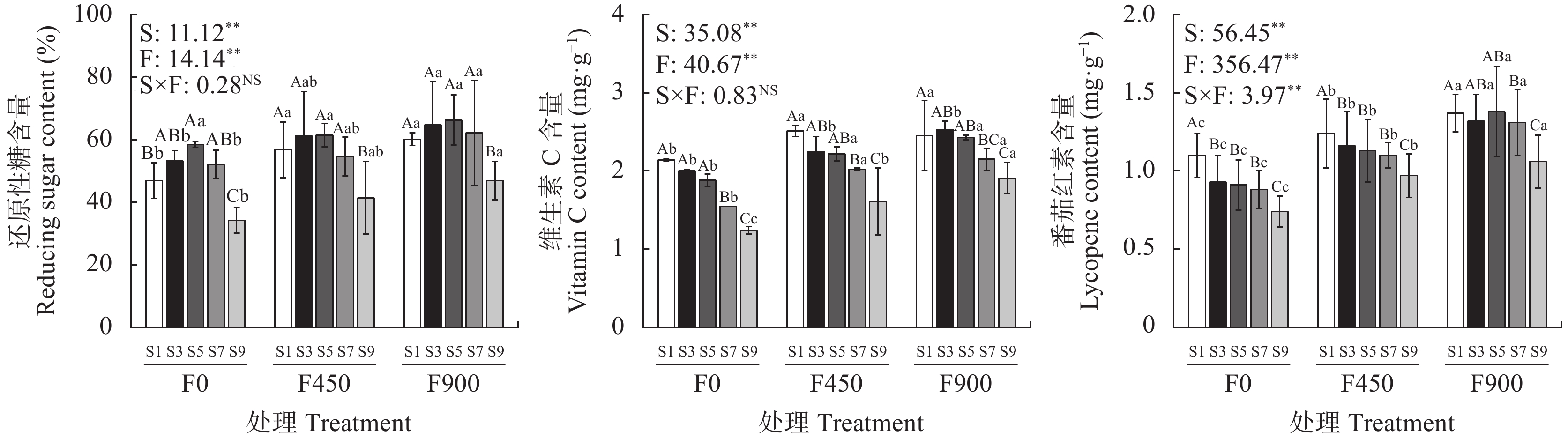
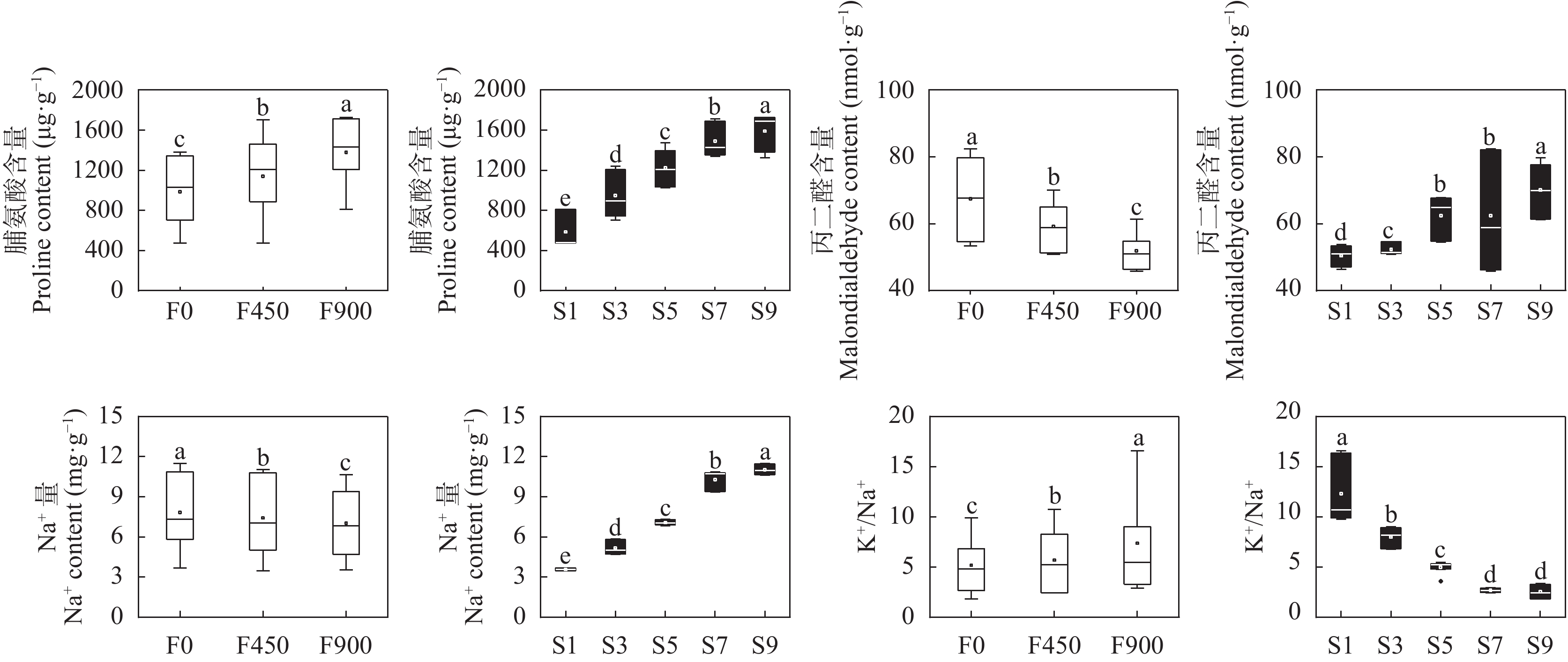
针对环渤海盐碱区淡水资源匮乏制约作物生长的问题, 依据区域咸水资源禀赋, 研究黄腐酸对咸水灌溉下番茄产量和品质的调控效应。采用基质栽培水肥一体化试验方法, 设置3个黄腐酸浓度水平: 0 mg·L−1、450 mg·L−1和900 mg·L−1; 5个咸水浓度水平: 1 g·L−1、3 g·L−1、5 g·L−1、7 g·L−1、9 g·L−1共15个处理。结果表明, 与不添加黄腐酸相比, 添加黄腐酸对不同浓度咸水灌溉下的番茄均有显著的增产效果(P<0.05), 添加450 mg·L−1、900 mg·L−1黄腐酸分别增产6.14%~21.08%和12.83%~34.63%。随着灌溉咸水浓度的增加, 番茄单果重、单株果实数目、耗水量、产量水分利用效率、果实维生素C和番茄红素含量显著下降, 果实还原性糖呈现先增加后下降的趋势; 施用450 mg·L−1和900 mg·L−1的黄腐酸均能提高咸水灌溉下番茄单果重、单株果实数目、耗水量、产量水分利用效率、果实维生素C、番茄红素、还原性糖含量。随着黄腐酸浓度的增加, 番茄叶片脯氨酸含量和K+/Na+显著增加, 丙二醛和Na+含量显著降低。单株产量和耗水量均与K+/Na+呈极显著正相关, 与脯氨酸量、丙二醛量、Na+含量呈极显著负相关; 番茄果实维生素C和番茄红素均与K+/Na+呈显著正相关, 与丙二醛量、Na+含量呈极显著负相关; 还原性糖与丙二醛量、Na+含量呈显著负相关。上述结果表明, 黄腐酸主要通过促进有机渗透调节物质脯氨酸积累、提高K+/Na+以及降低膜脂过氧化产物丙二醛的产生缓解咸水灌溉对番茄产量的抑制, 同时还能提高产量水分利用效率、果实维生素C、番茄红素和还原性糖含量, 改善番茄品质。
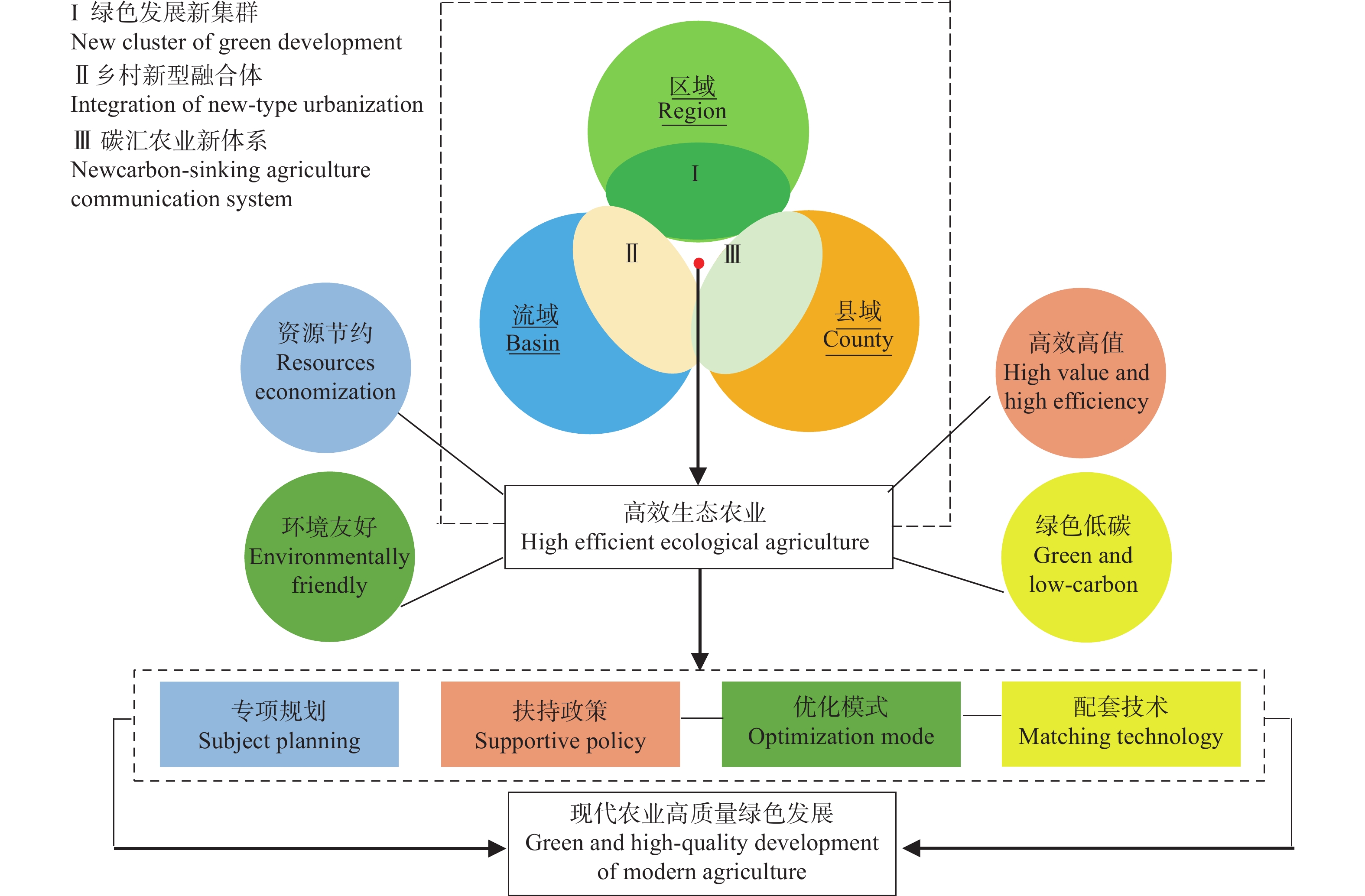
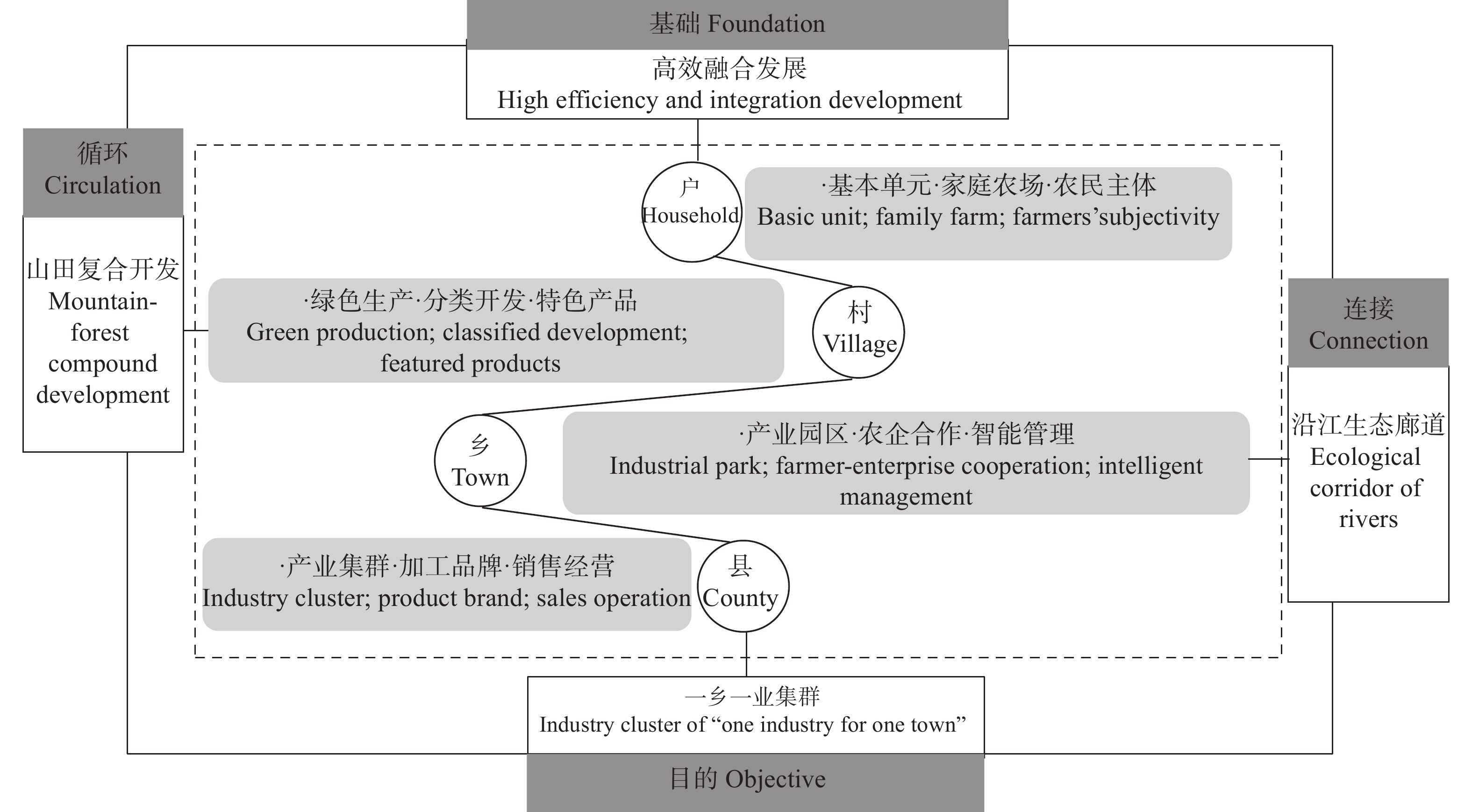
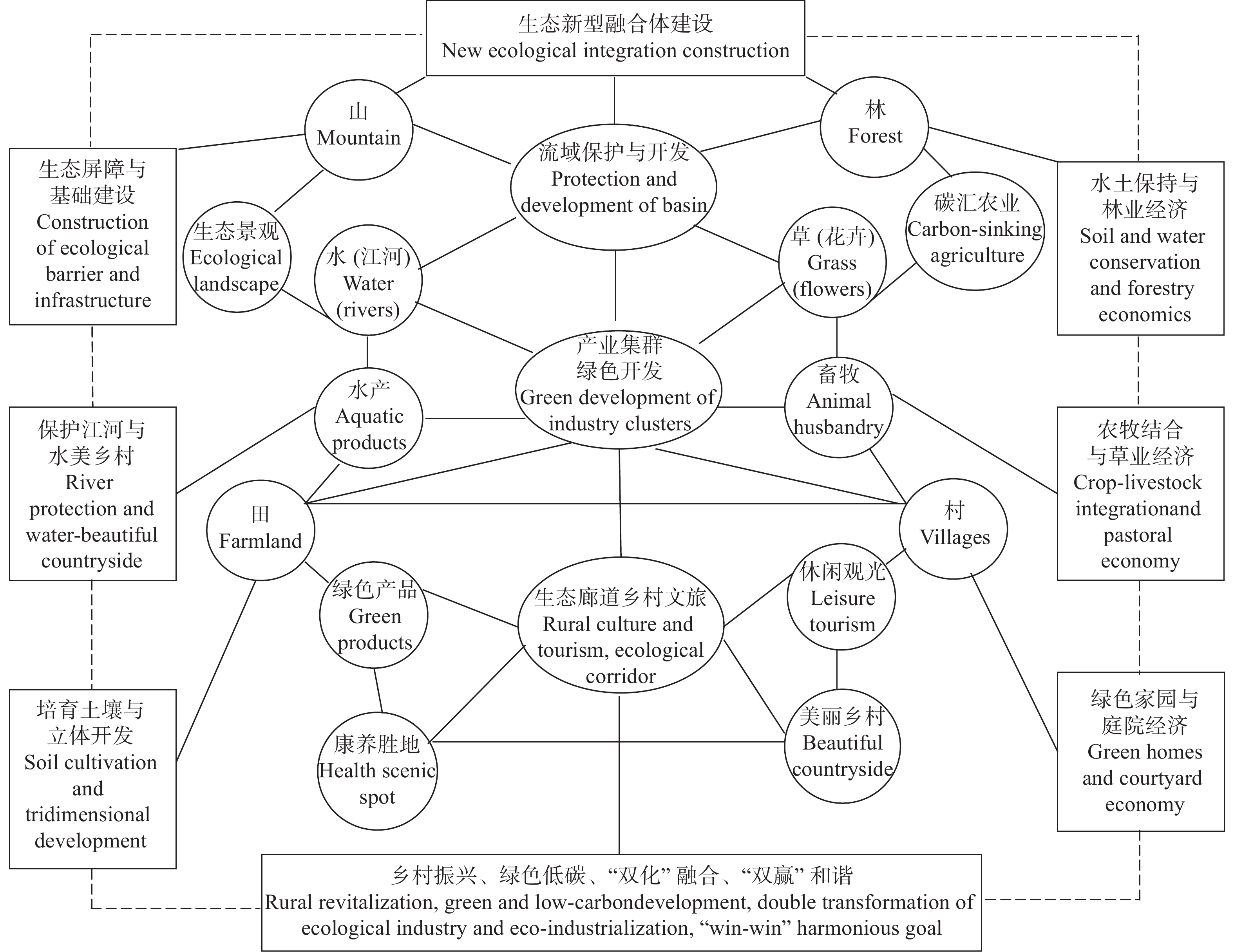
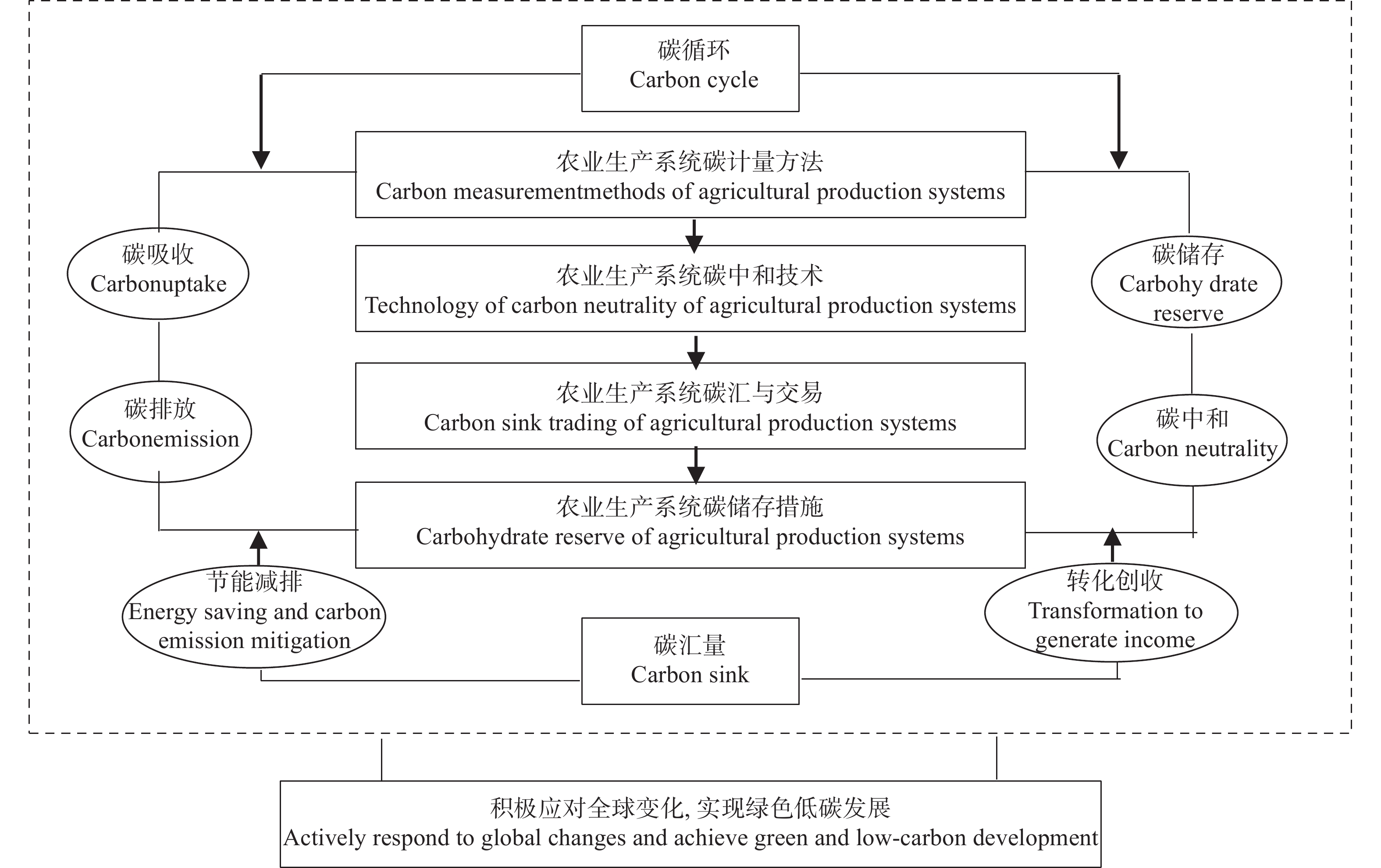
高效生态农业的应运兴起丰富了农业强国建设的内容, 对其深入探索与不断完善是实施乡村振兴战略的重要举措与运营载体。本研究在总结保障供应、资源节约与环境友好型农业发展经验的基础上, 进行高效生态农业的科学内涵和理论内涵的延伸和拓展, 构建“县域-区域-流域”高效生态农业建设主体框架, 剖析高效生态农业在乡村振兴进程中的建设思路和发展目标。高效生态农业是生态农业的提高和升华, 本研究依照福建省现代农业提质增效与农业强省的建设要求, 提出福建乡村生态产业化与产业生态化融合发展新集群构建; 因地制宜营造人与自然和谐共生的宜业宜居乡村融合体; 深入探索并创立福建乡村碳中和管理新机制与创业新体系等新技术体系优化构建与实施对策, 进而探索创建乡村产业化+生态化“双化协同”开发与农业经济效益+生态效益“双益统筹”发展的生产经营模式, 因地制宜推动富有区域特色的高效生态农业转型升级, 是全面推进乡村振兴的有效路径。
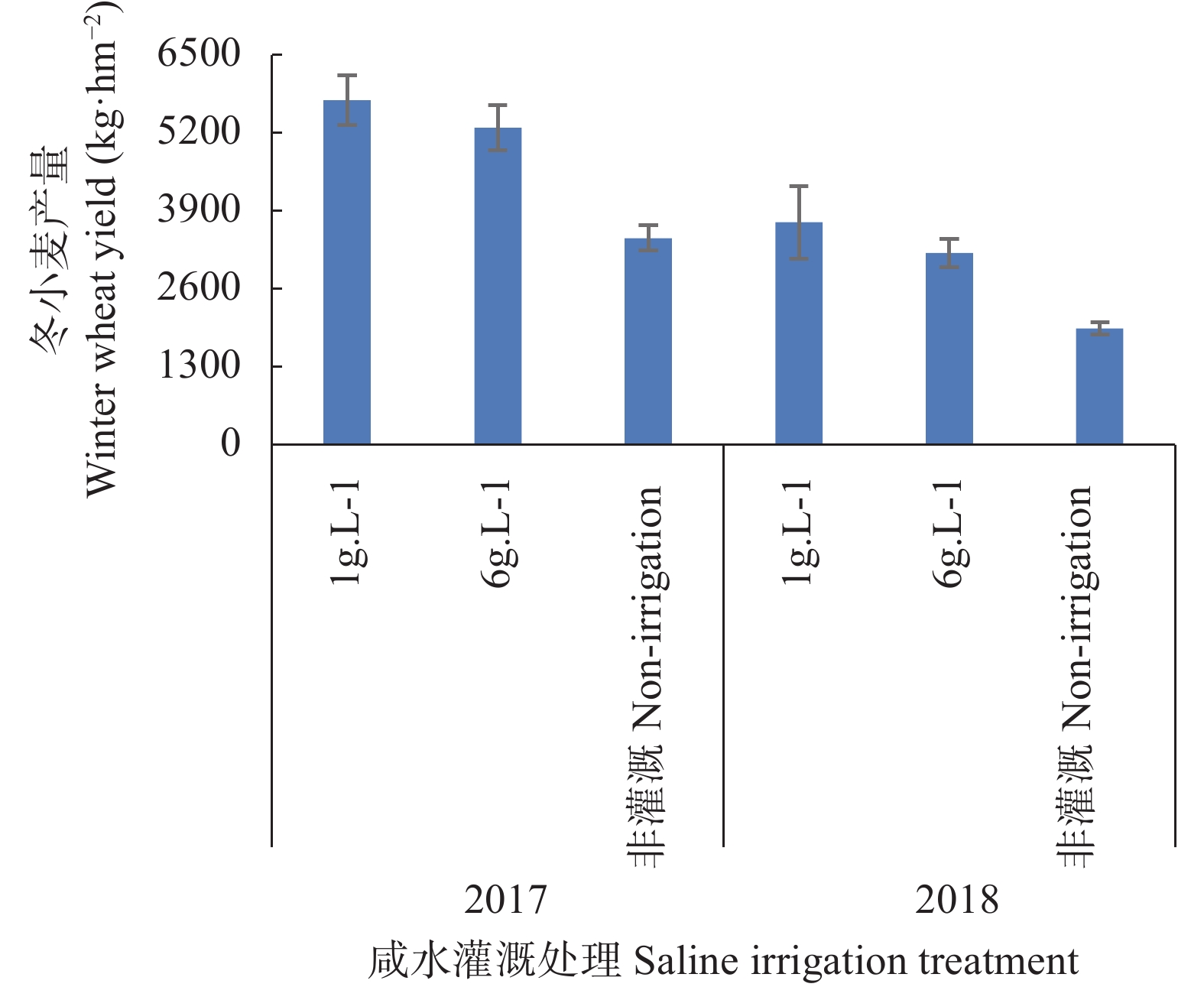
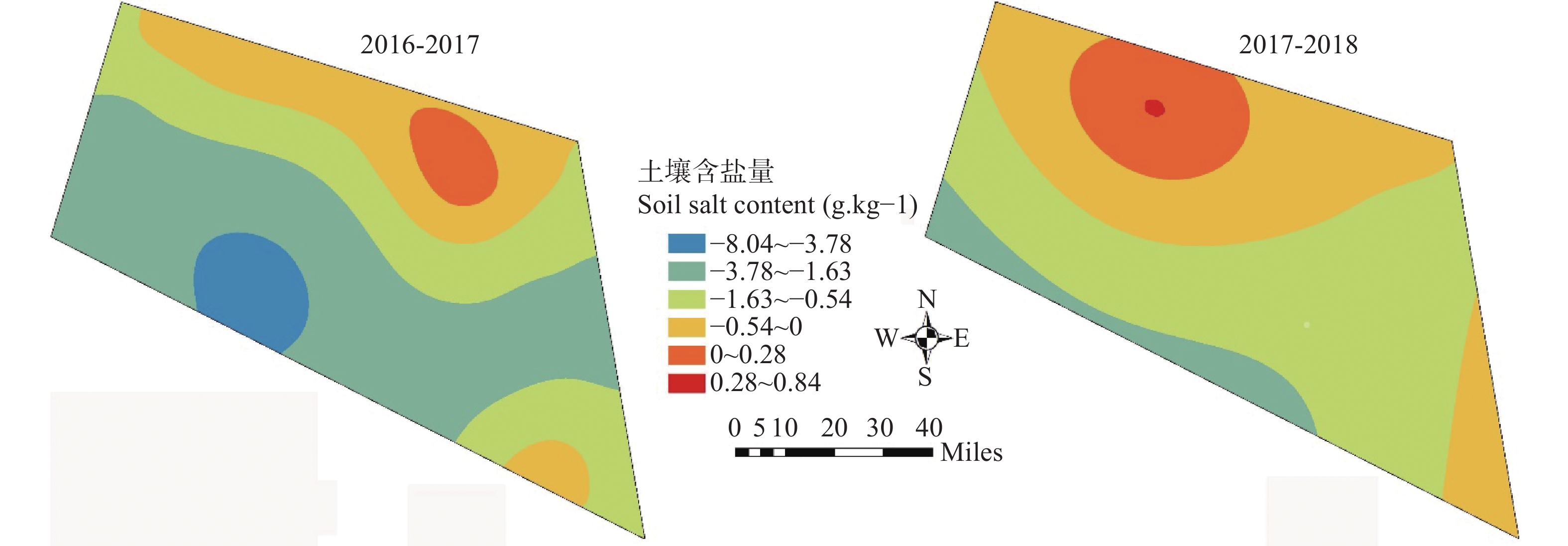
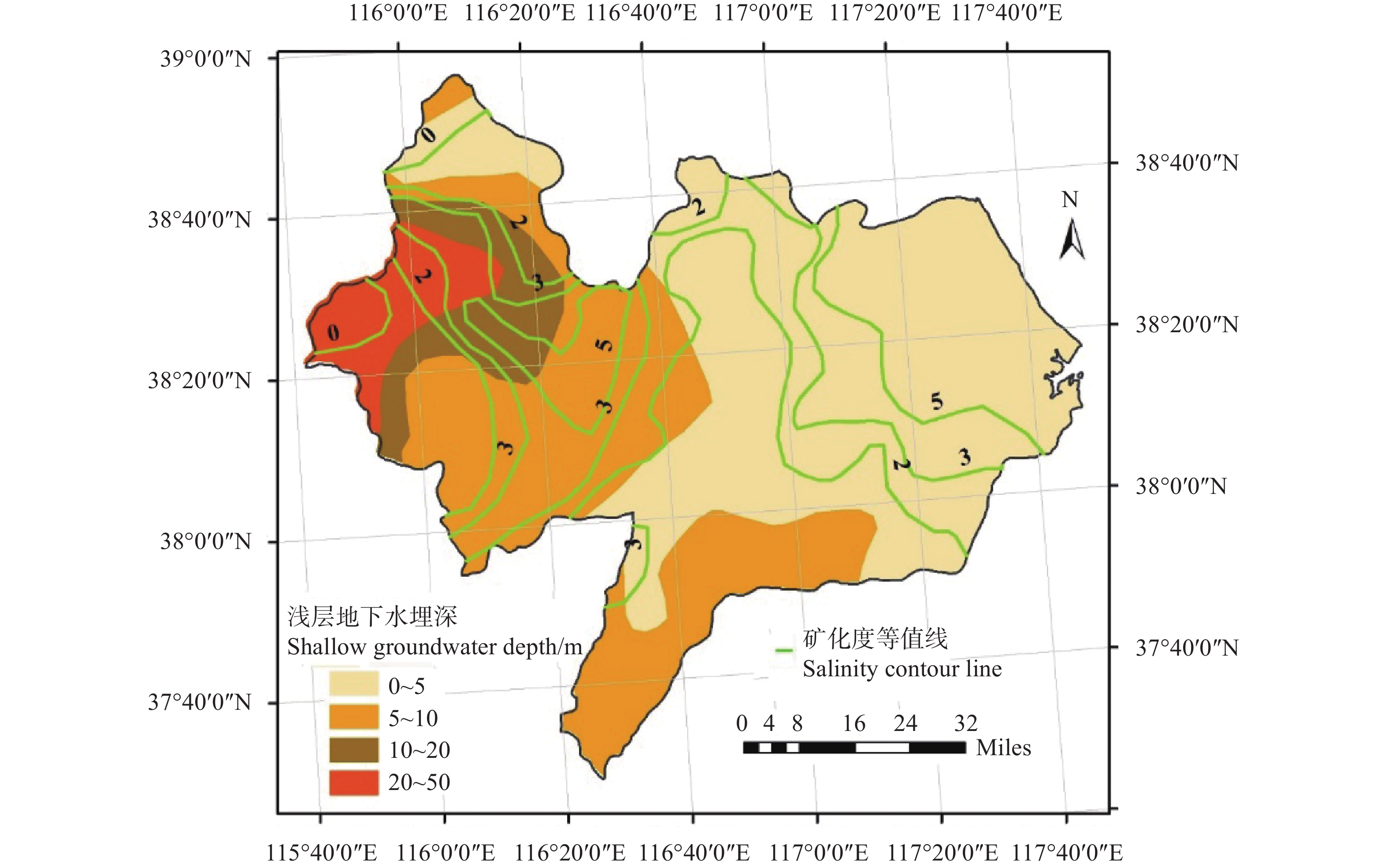
河北滨海盐碱耕地缺少灌溉水资源, 该区具有丰富的浅层微/轻度咸水资源, 冬小麦生长关键期适当进行微/轻度咸水灌溉能够缓解该区农田灌溉危机。本文以河北省滨海盐碱地为研究对象, 选择沧州市南大港产业园区马营村为试验基地, 开展浅层轻度咸水冬小麦农田灌溉试验, 分析咸水灌溉对冬小麦产量和土壤盐分含量变化的影响。结果表明: 1)河北滨海盐碱地浅层轻度咸水(6 g∙L−1)在冬小麦拔节—抽穗期进行一次40 m3的灌溉, 产量能够达到旱作冬小麦的1.5倍以上, 增产效果明显。2)基于暗管排水排盐工程、在雨季保持多年平均降雨量条件下, 2016—2018年雨季后实验地土壤平均含盐量分别为2.86 g∙kg−1、1.60 g∙kg−1和1.38 g∙kg−1, 呈现逐年降低的趋势。由此得出研究结论: 河北滨海盐碱地浅层咸水资源在冬小麦灌溉方面能够实现安全利用。在此基础上, 以河北滨海沧州市为例分析浅层咸水资源前景表明, 从水资源量上, 1~5 g∙L−1的浅层微/轻度咸水资源可以满足冬小麦农田因地下水压采限采造成的至少1.7×108 m3灌溉水量亏缺, 沧州市耕地可以扩大一年两熟种植面积, 增加冬小麦种植面积约9000 hm2。以上研究结果可为河北滨海盐碱地咸水资源安全利用、土壤健康和粮食安全提供科学依据。

发展缓控释肥料是实现化肥减量增效的重要方向, 在支撑现代农业绿色发展中发挥着重要作用。文章回顾了北京市农林科学院植物营养与资源环境研究所在该领域32年的研究工作, 从研究历程、队伍建设、产品开发和农化服务等方面阐述了缓控释肥料自实验室走向农田的来龙去脉, 对未来的研究方向进行了分析与展望。研究所1991—1998年相继研制出沸石包膜肥料和聚烯烃树脂包膜肥料。1999—2015年,控释肥料逐步实现产业化。2016年以后,生物基包膜控释肥料受到重视,研究取得了较大进展。沸石包衣肥料是以天然沸石为包膜剂的无机包膜肥料。聚烯烃树脂包膜肥料是将树脂溶液喷洒在肥料表面形成一层半渗透或不渗透材料,以实现养分的控制释放。聚氨酯包膜肥料是一种热固性树脂包膜控释肥料,无溶剂原位反应成膜是其常用生产工艺,研究所先后开发了半自动、自动化连续生产技术。研究所缓控释肥料的创制服务了从大田到园艺系列作物养分精准需求, 带动了肥料产业升级、面源污染治理、肥料减量增效工作的开展。为适应农业发展的现实需求, 未来仍需在肥料产品包膜材料生物可降解、肥料养分释放多阶段连续可控、大规模连续化生产工艺创新、肥料产品质量在线快速检测技术、作物专用多元复合控释功能肥料等方面持续研究。
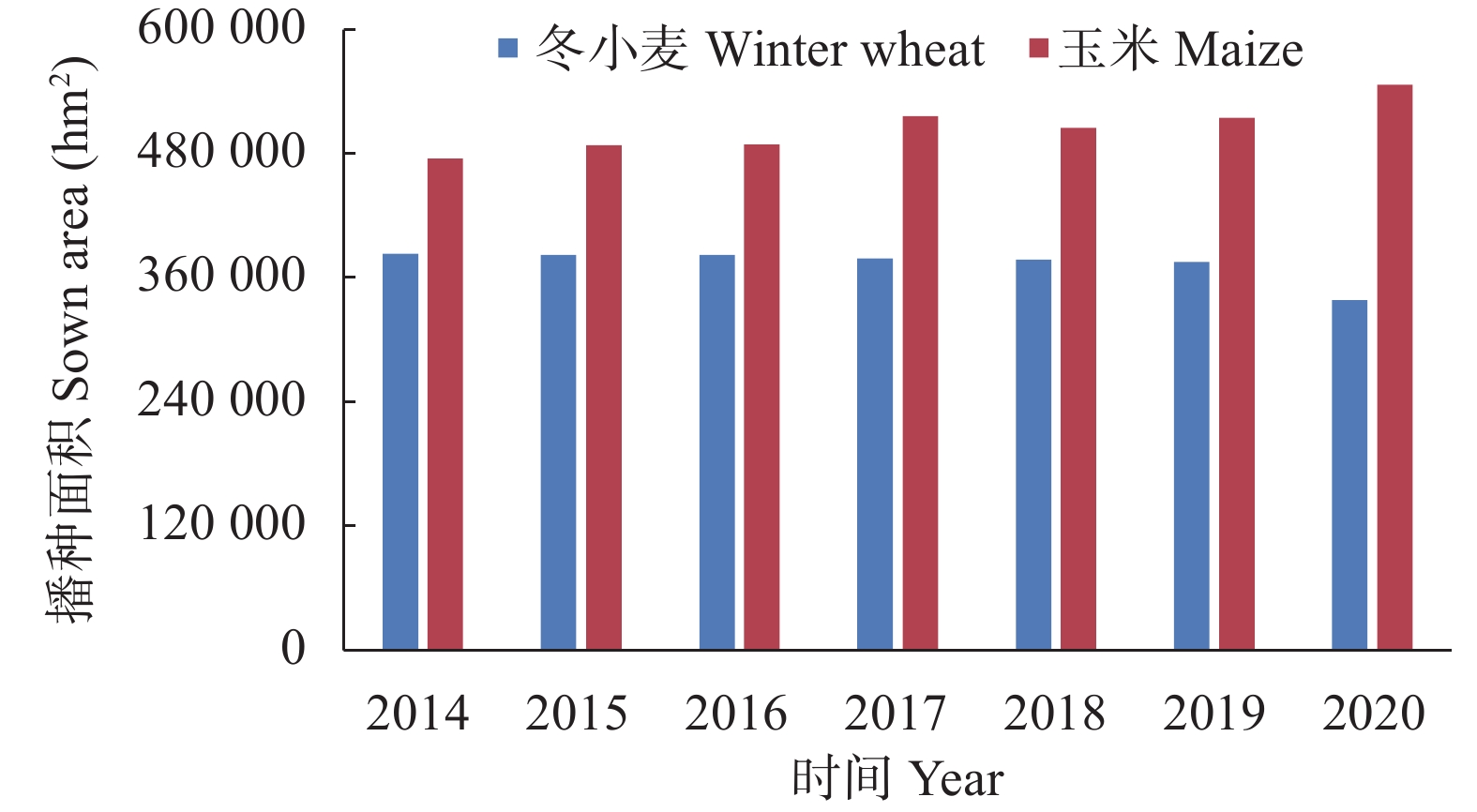
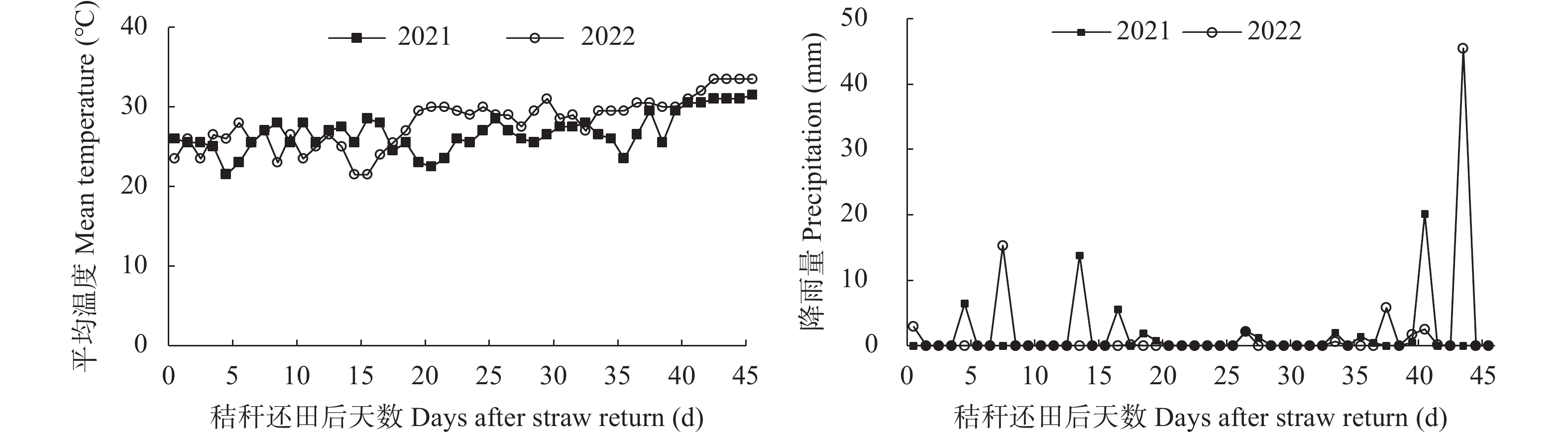
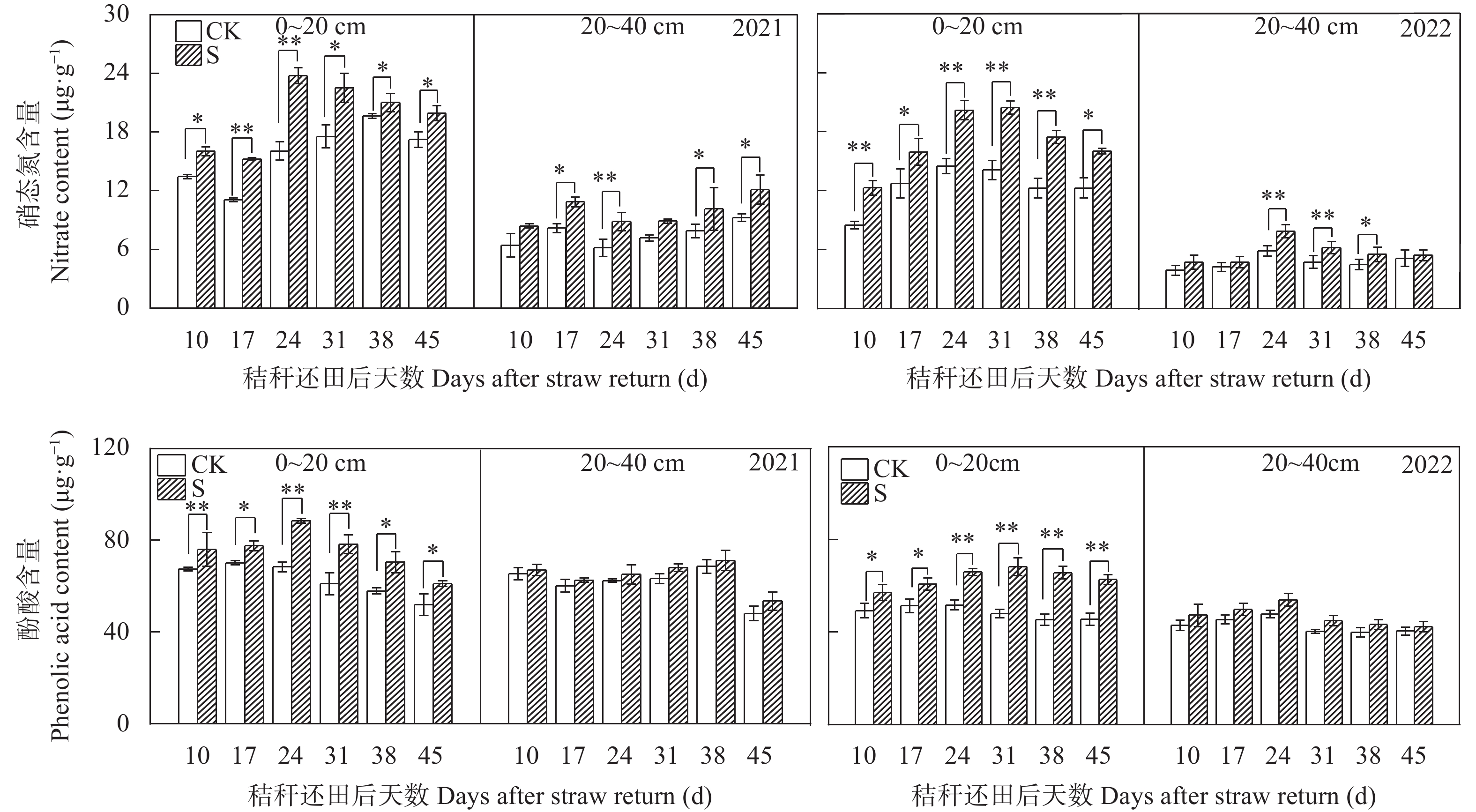
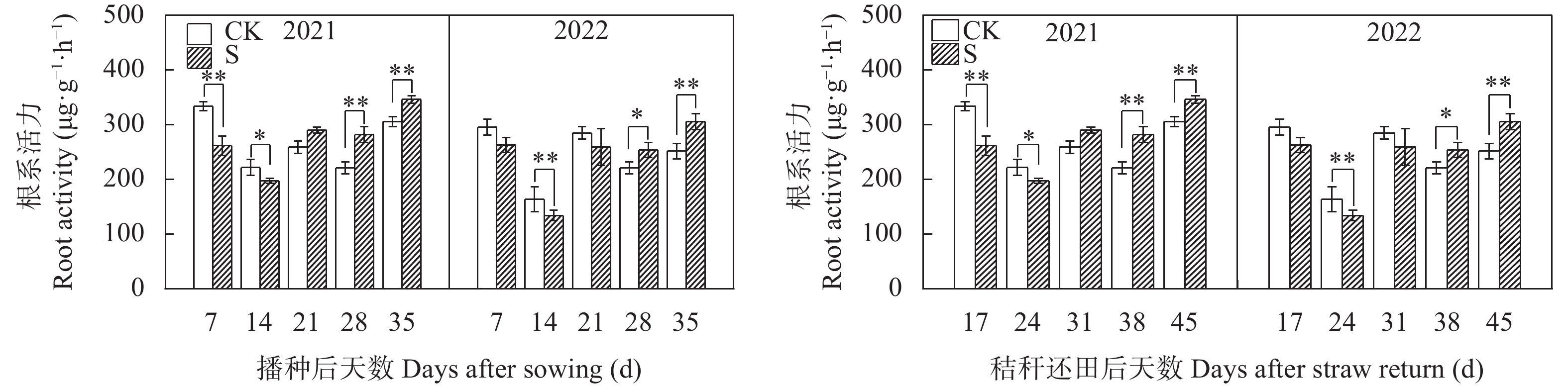
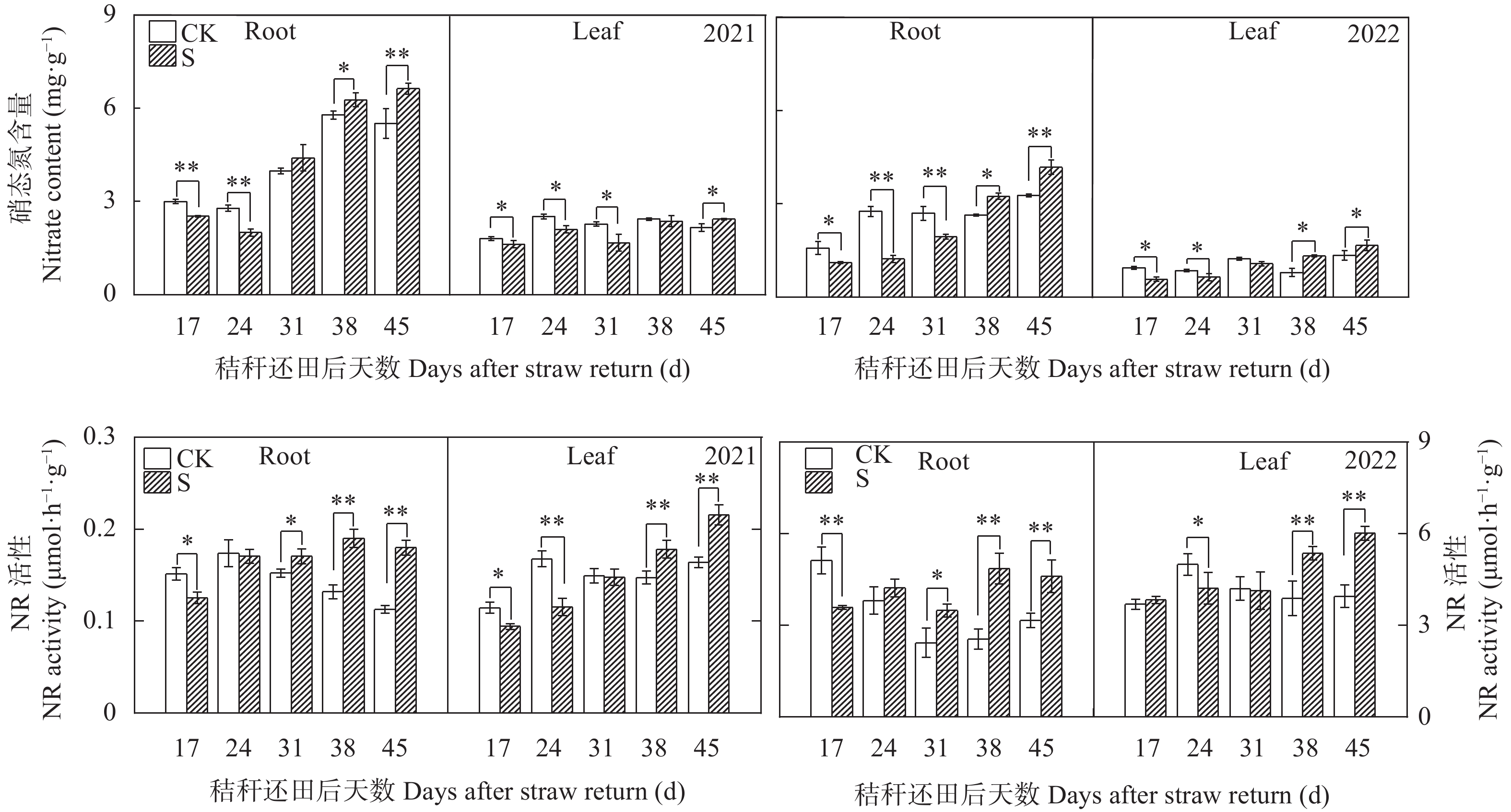
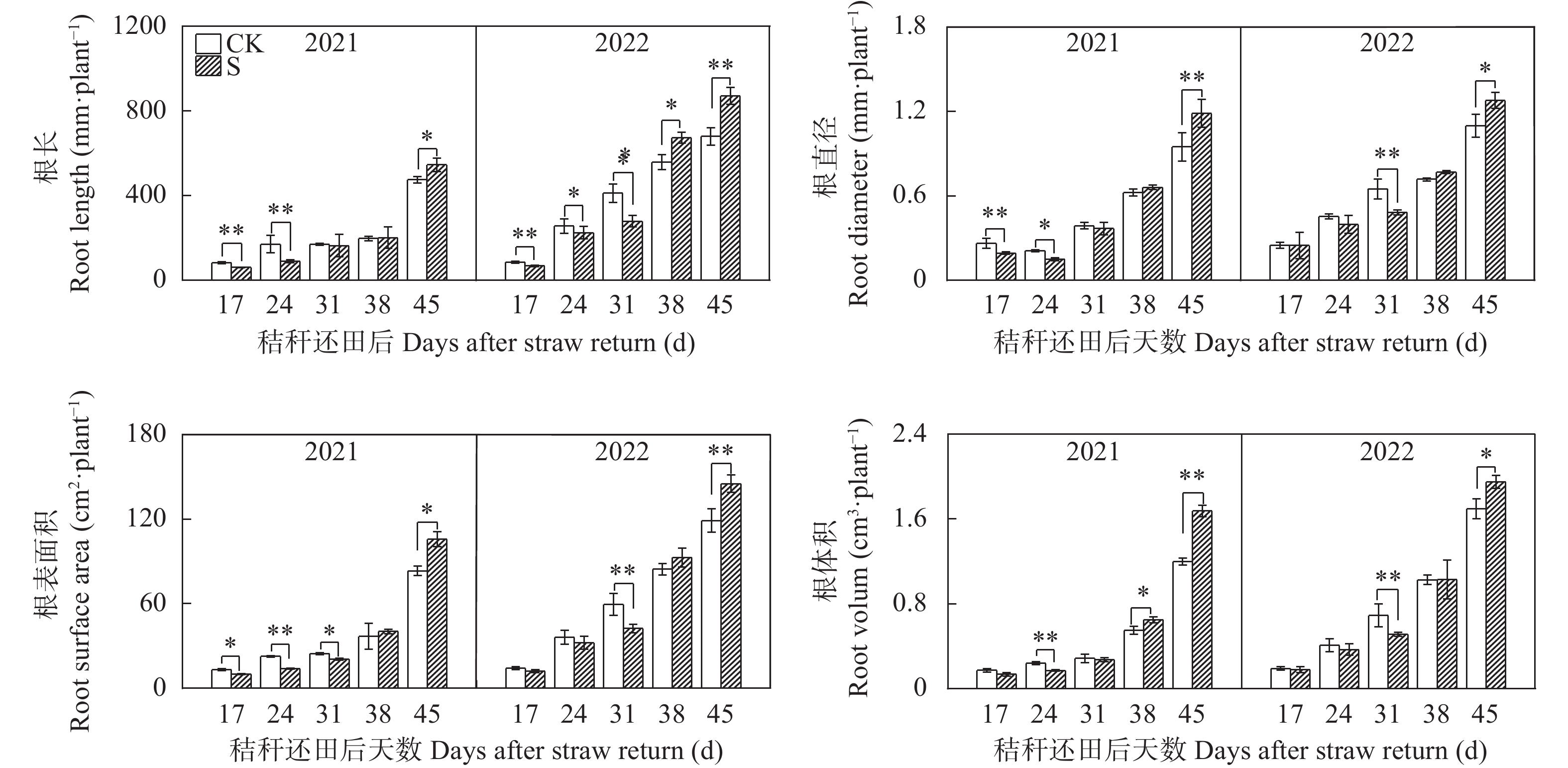
揭示前茬小麦长期秸秆还田后后茬棉田土壤硝态氮(NO3−-N)与酚酸含量的时空变化影响棉苗根系生长的生理机制, 可为完善秸秆还田技术提供理论支撑。基于长期小麦秸秆定位还田, 试验于2021年与2022年进行。以棉花品种‘中棉所425’为材料, 设置小麦秸秆不还田(CK)与小麦秸秆还田(S) 2个处理, 研究长期小麦秸秆还田下后茬棉田土壤NO3−-N和酚酸含量的时空变化对棉苗根系生长与NO3−-N吸收的影响。结果表明, 秸秆还田增加了土壤NO3−-N与酚酸含量, 对0~20 cm土层的影响大于20~40 cm;随秸秆还田时间推迟, 土壤NO3−-N与酚酸含量呈先增加后降低趋势, 且在秸秆还田后24~31 d达到峰值。秸秆还田后31 d前, 秸秆还田处理棉株根系活力、根系NO3−-N含量、硝酸还原酶活性、根系生物量和形态指标均显著低于CK处理, 31 d后则呈相反趋势。相关分析表明, 0~20 cm土壤酚酸浓度与根系活力、根系NO3−-N含量、棉花根系长度、直径、表面积和地上部生物量呈显著负相关; 不同土层NO3−-N含量与棉苗形态、生理指标及生物量之间呈正相关但未达显著水平。秸秆还田对棉花幼苗生长影响呈“先抑后促”的趋势, 秸秆还田后31 d前, 酚酸含量的增加降低了棉苗根系活力和根系生长、抑制了棉苗对NO3−-N的吸收利用, 表明秸秆还田前期对棉株生长的“抑制效应”大于秸秆的“肥料效应”, 秸秆还田31 d后, 秸秆的“肥料效应”大于酚酸的“抑制”效应, 促进棉株根系的生长。
中国农业正处于现代化转型的关键时期, 进一步明确新时期中国生态农业现代化的内涵、任务和路径, 并达成广泛共识十分必要。现代生态农业源于传统农业, 吸纳现代科技成果, 具备现代产业经营体系, 是中国式农业现代化的根本方向。中国生态农业现代化面临保障食物安全、生态安全和营养安全, 传承农耕文化, 支撑共同富裕的艰巨任务与挑战。今后应坚持系统观念和守正创新原则, 从创新生态农业典型模式、加强农业绿色投入品研发、加快生态农业新装备的研发与应用、提升生态农业产业链运营能力和完善生态农业生态补偿政策与机制等方面重点突破。依靠科技与改革双轮驱动, 创新发展适合我国国情的现代生态农业产业技术体系, 为世界农业可持续发展提供中国解决方案。
乡村是国土空间的重要组成部分,乡村减排增汇是实现双碳目标的关键举措。本文基于人地关系地域系统理论构建乡村碳循环体系,并针对乡村生产和生活活动,从核算方法、量化结果、影响因素与治理途径四方面综述当前国内外定量研究成果,以期形成乡村空间碳效应的系统认知。结果表明:①乡村碳源主要为住房、交通和烹饪等生活活动和粮食作物种植等农业生产活动,生活碳排放约为农业生产碳排放的2倍。以5%的清洁能源代替煤炭消耗有望减少60%的乡村生活碳排放。②乡村碳汇主要依靠作物生长发育等自然过程,我国乡村农田土壤年均固碳量约0.25亿吨,可抵消农业碳排放总量的12.68%。通过农田管理,农田土壤碳汇每年可抵消农业碳排放的61.26%。③在保障粮食安全的前提下,乡村碳减排应从乡村数字化能源系统建设、清洁能源技术研发、农业废弃物循环利用三方面入手,提高能源利用效率,改善能源结构。乡村碳增汇需着力加大农田管理力度,扩展秸秆粉碎翻压还田、少免耕、施用生物炭等保护性耕作措施应用规模,强化农田土壤碳汇能力。研究可为优化乡村环境治理体系,改进乡村治理策略,推动乡村低碳转型和高质量发展提供决策参考。
三叶青作为“新浙八味”之一,具有抗肿瘤、抗病毒和退热等临床效果,主要活性成分为黄酮类化合物。三叶青生长在林间荫蔽环境,其活性成分合成、生长发育、生理变化等均会受到林下短波光调控。本文为明晰短波光对三叶青品质及生理变化的调控规律,选择了蓝光和紫光两种短波单色光作为处理光源,以白光作为对照,处理7 d、15 d、30 d、45 d和60 d,测定三叶青块根和叶片中总黄酮含量、黄酮代谢合成酶活性、叶片生理生化指标(抗逆酶活性和丙二醛、初生代谢可溶物、光合色素含量),以期探讨三叶青黄酮合成过程和生理生化变化对短波光条件的时空响应及规律。结果表明,短波单色光处理可使三叶青处于轻度胁迫状态,蓝光处理后三叶青中总黄酮含量最高(叶片137.75 mg/g ,块根149.00 mg/g );紫光处理使叶片和块根中的PAL酶活性(96.50 U/g FW、109.47 U/g FW)和叶片中CHI酶活性(65.17 U/g FW)达到最高,蓝光处理使块根中的CHS酶活性(46.76 U/g FW)达到最高。此外,光合色素和抗逆能力和可溶性物质对短波光的响应度有差异,与白光相比,蓝光和紫光均可促进渗透调节物质的积累。本研究的结果可为光调控三叶青品质提供理论基础和实践指导。
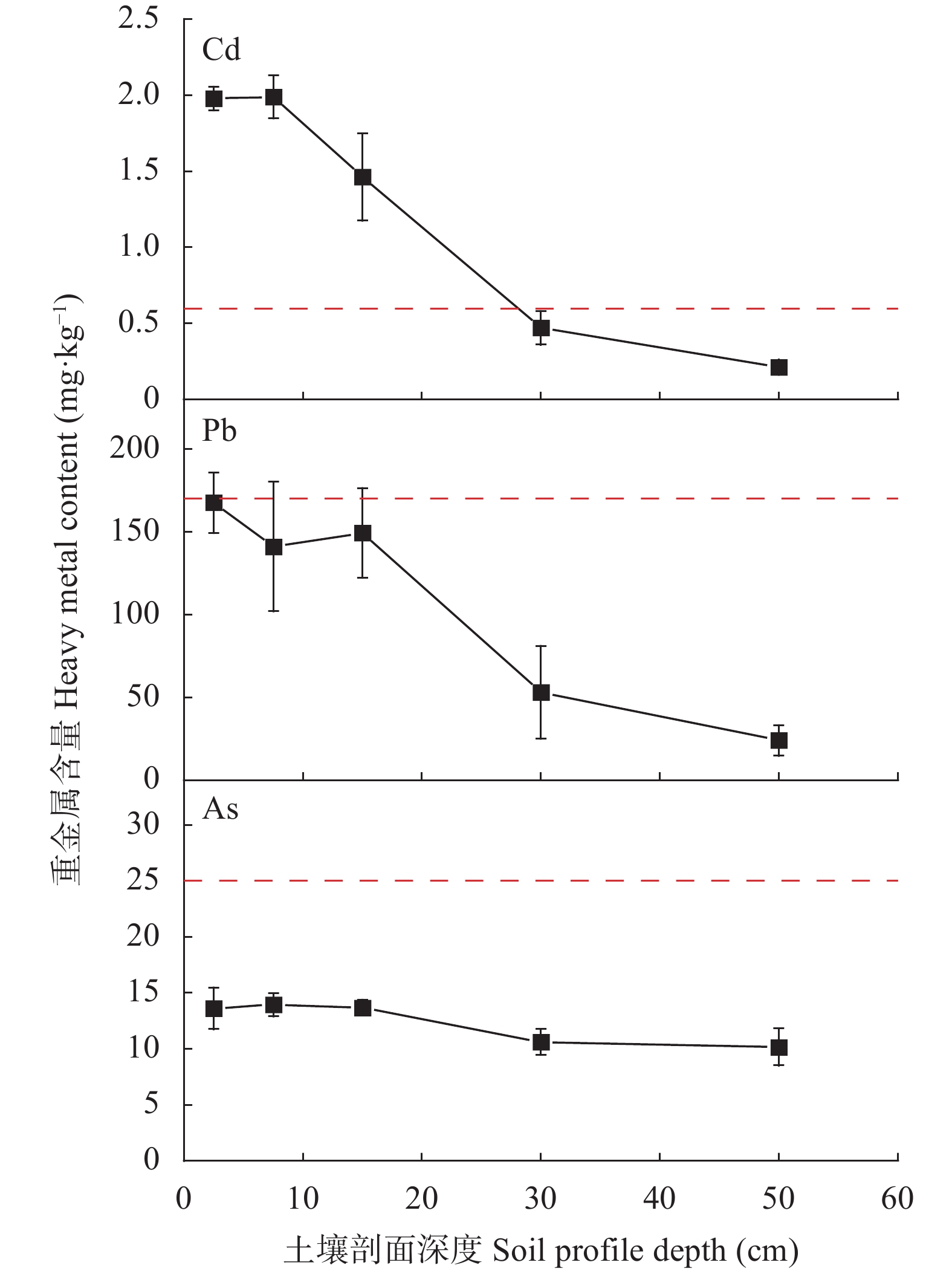
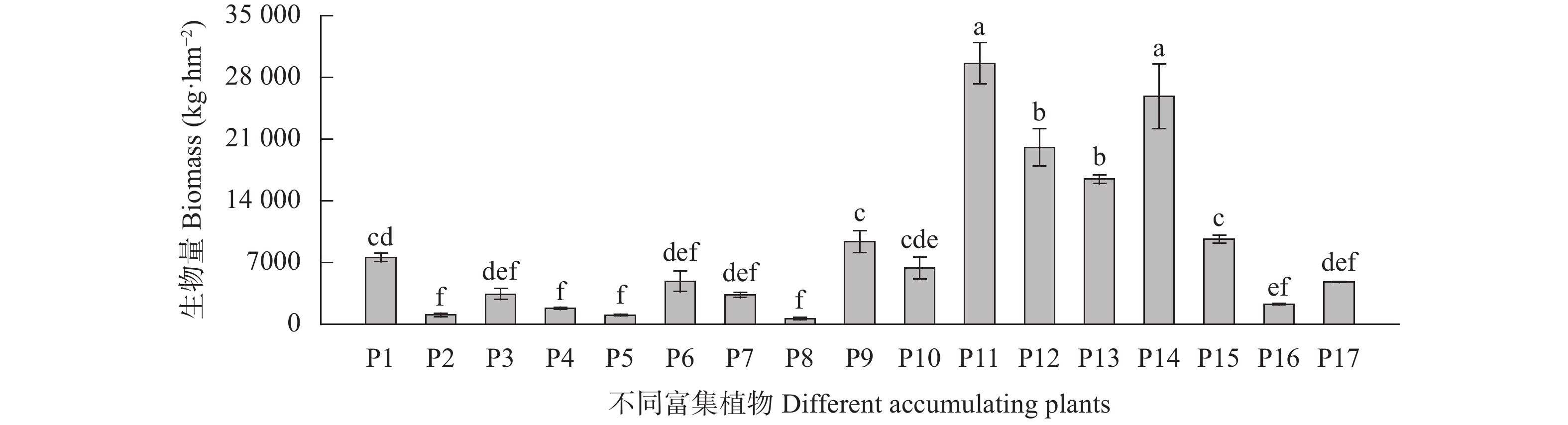
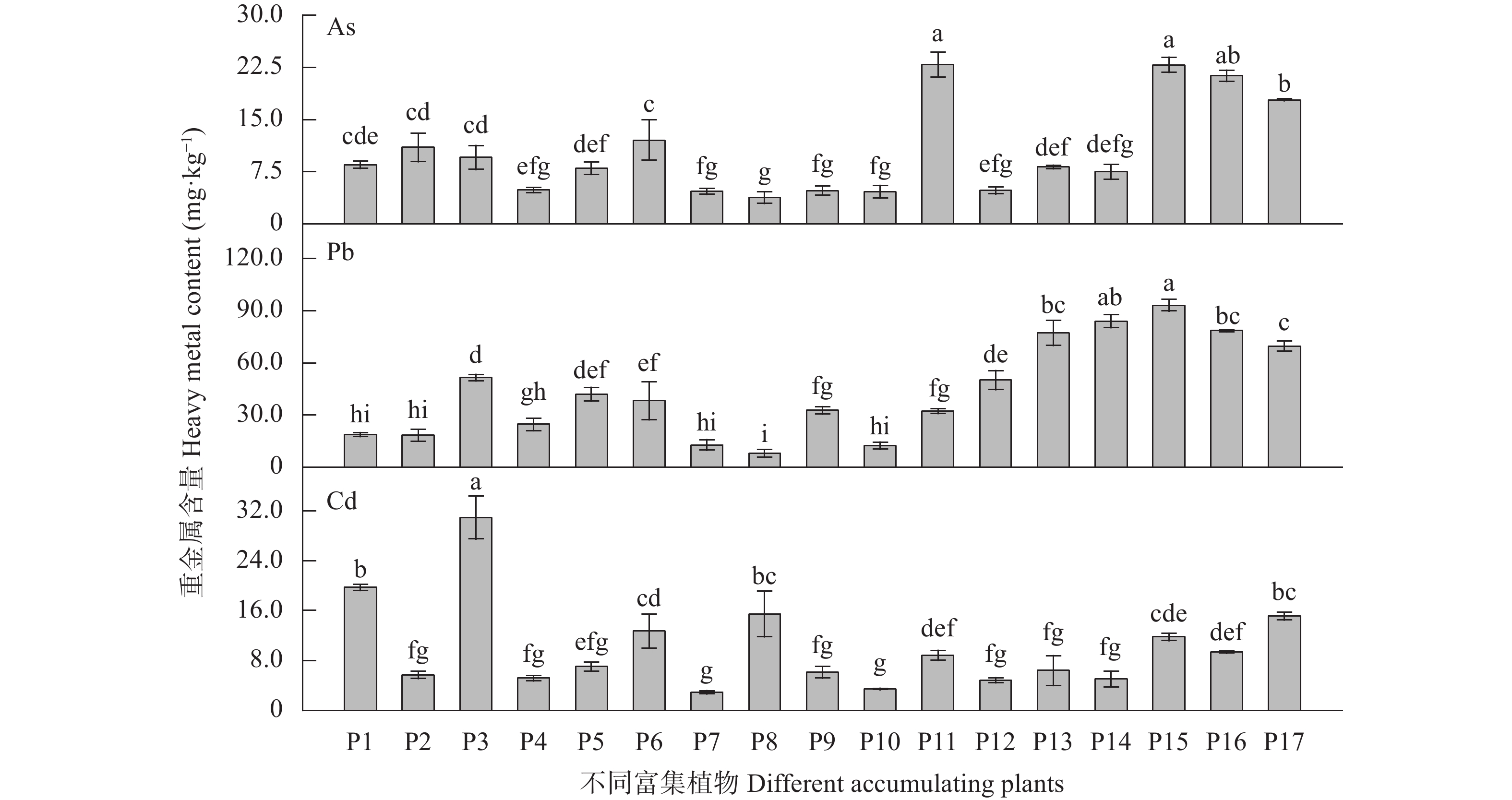
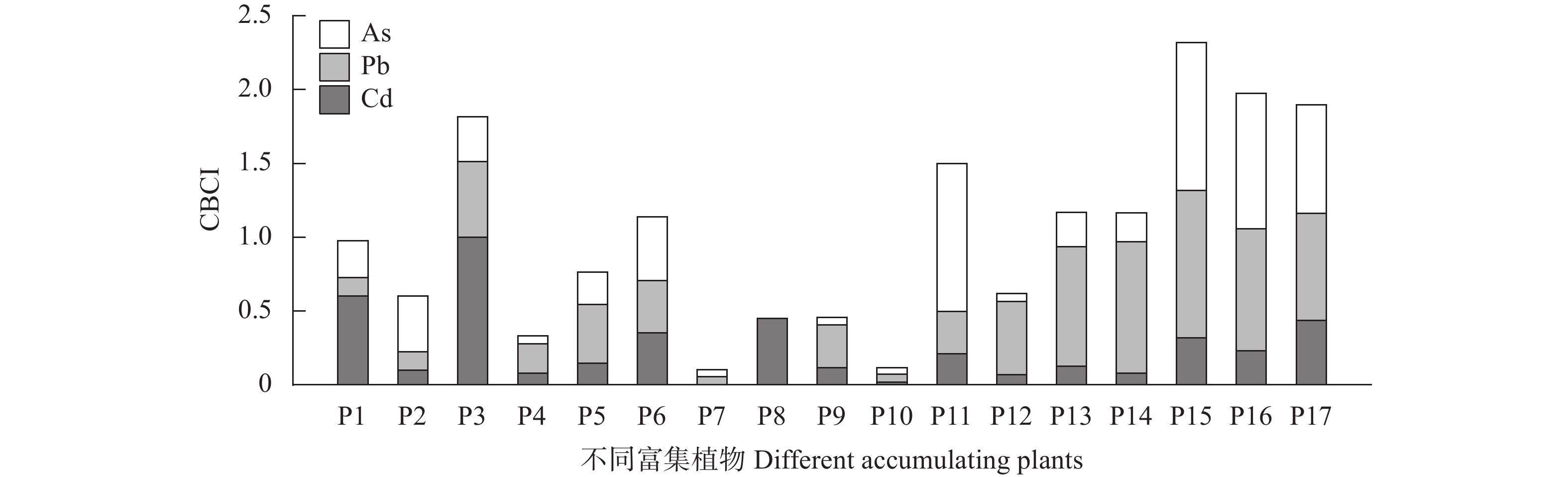
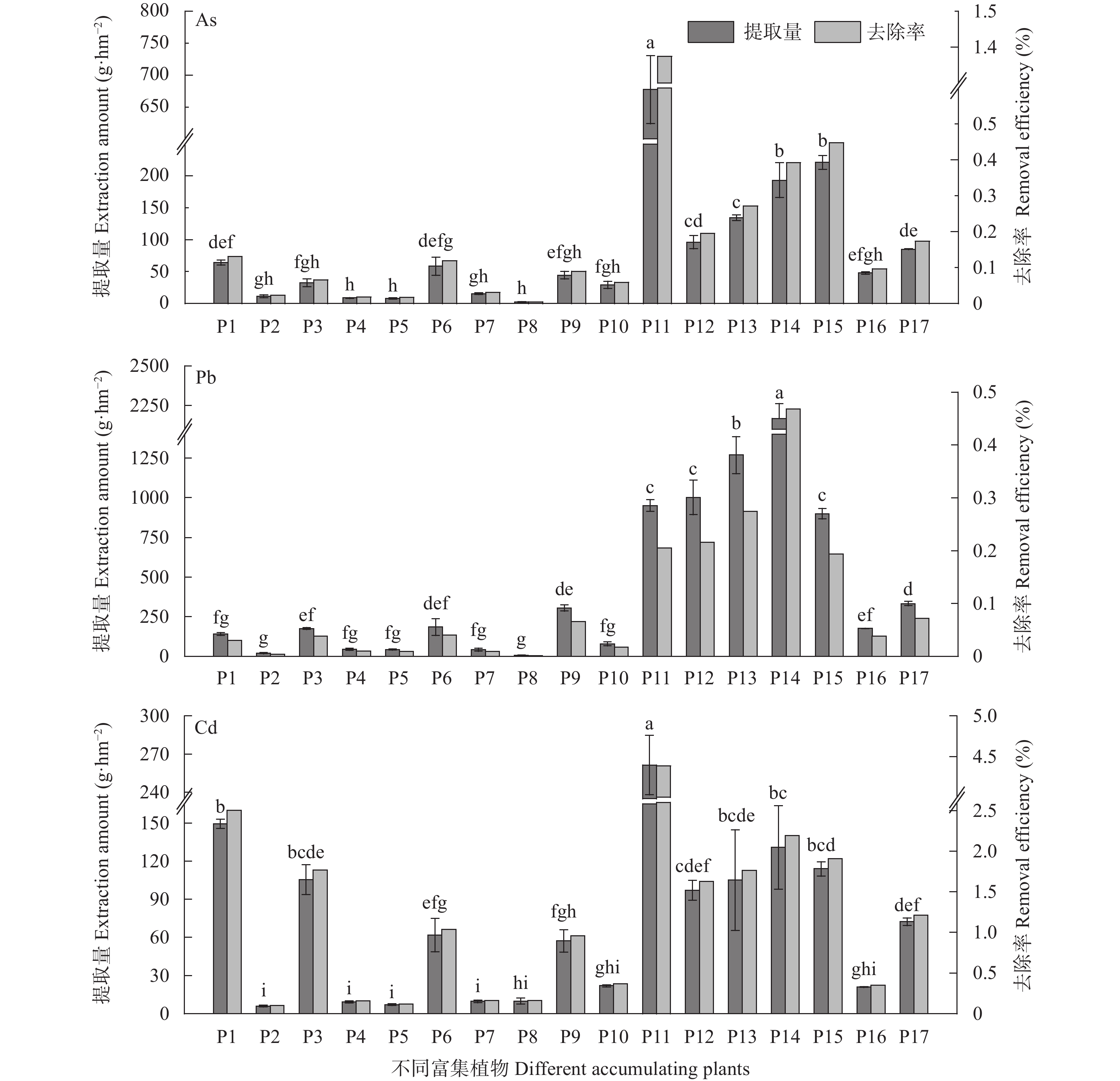
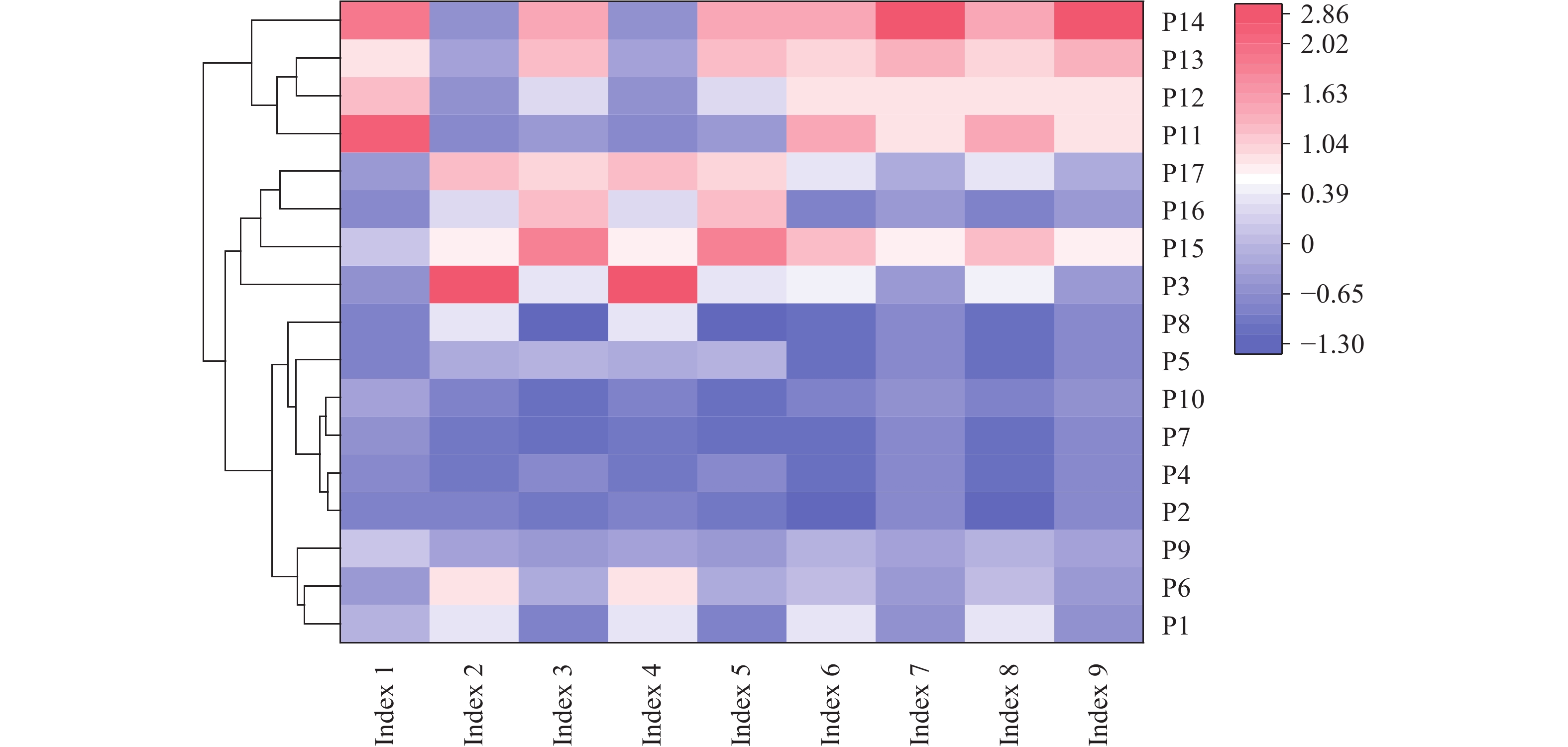
为探究有色金属企业周边农田重金属污染状况, 筛选修复与安全利用优势植物。本研究在河南省济源市某铅锌冶炼企业周边农田开展调查, 探明研究区土壤重金属污染特征, 并选用17种重金属富集植物, 开展田间试验, 比较其生物量、重金属含量、富集系数、提取量及去除率等, 建立适合研究区的土壤重金属污染植物修复技术模式。结果表明, 试验区土壤镉(Cd)、铅(Pb)存在超标情况, 超标率Cd>Pb, 污染物主要分布在0~20 cm耕作层, 属典型的轻中度Cd、Pb污染农田土壤。试验区主要农产品小麦籽粒Cd、Pb超标率为100%, 部分样品As超标, 玉米籽粒符合标准。17种富集植物生物量和对重金属的富集能力差异较大, 籽粒苋生物量最高, 达29 598 kg∙hm−2, 是遏蓝菜的46.6倍, 植物体内重金属Cd、Pb、As含量分别为2.90~30.90 mg∙kg−1、7.81~93.1 mg∙kg−1、3.76~22.9 mg∙kg−1。不同植物对土壤中重金属的去除率存在明显差异, 籽粒苋、冬油菜‘中油千斤旱’、向日葵‘S606’表现出良好的Cd、Pb提取修复潜力, 对土壤中Cd的去除率均高于1.90%。此外, 油菜和向日葵可进一步加工生产食用油、饲料或有机肥料, 实现边修复边生产, 应用前景较好。在研究区采用籽粒苋—冬油菜轮作和向日葵单作两种修复模式均能达到较高的修复效果, 具备一定的推广应用潜力。
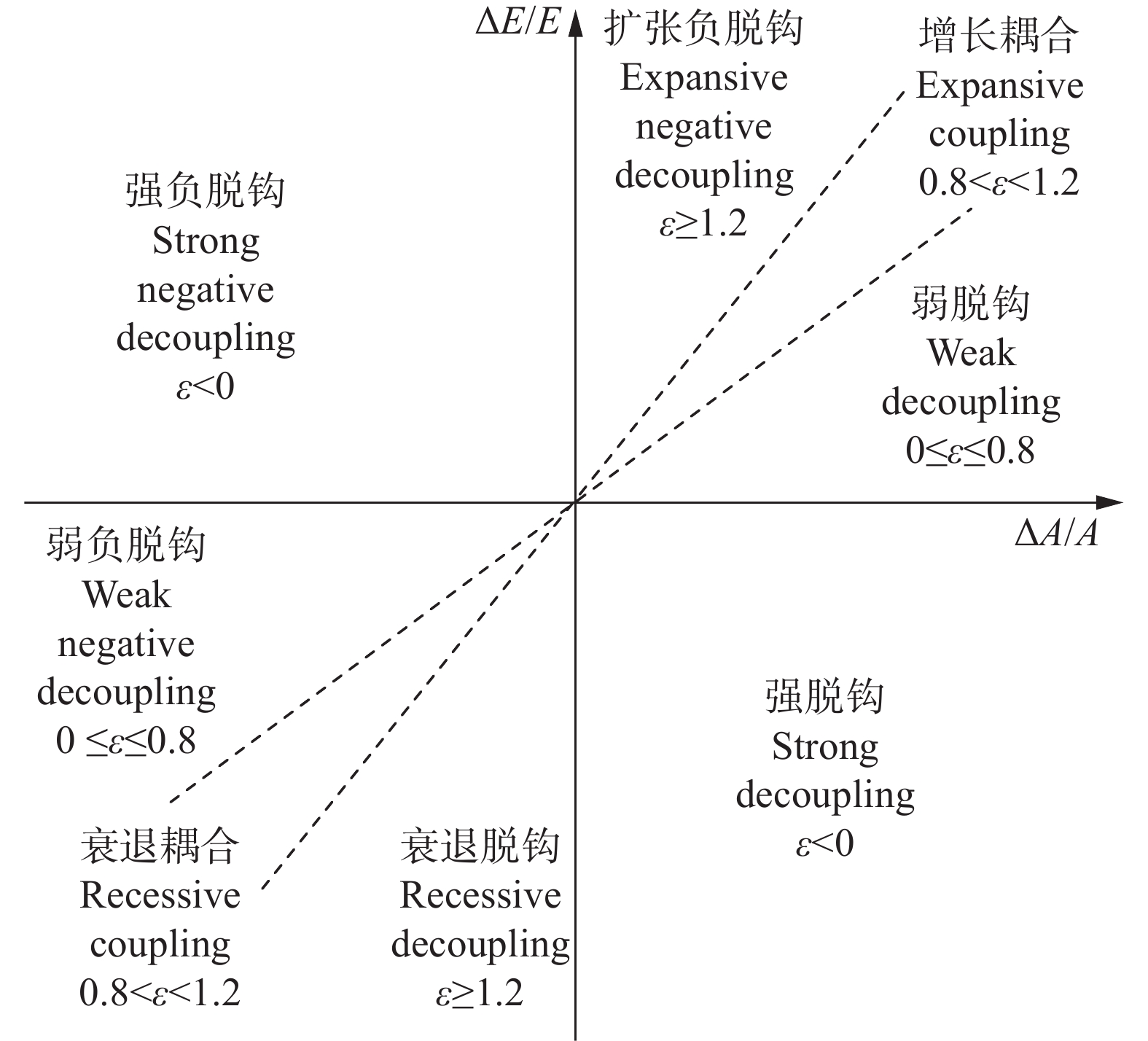
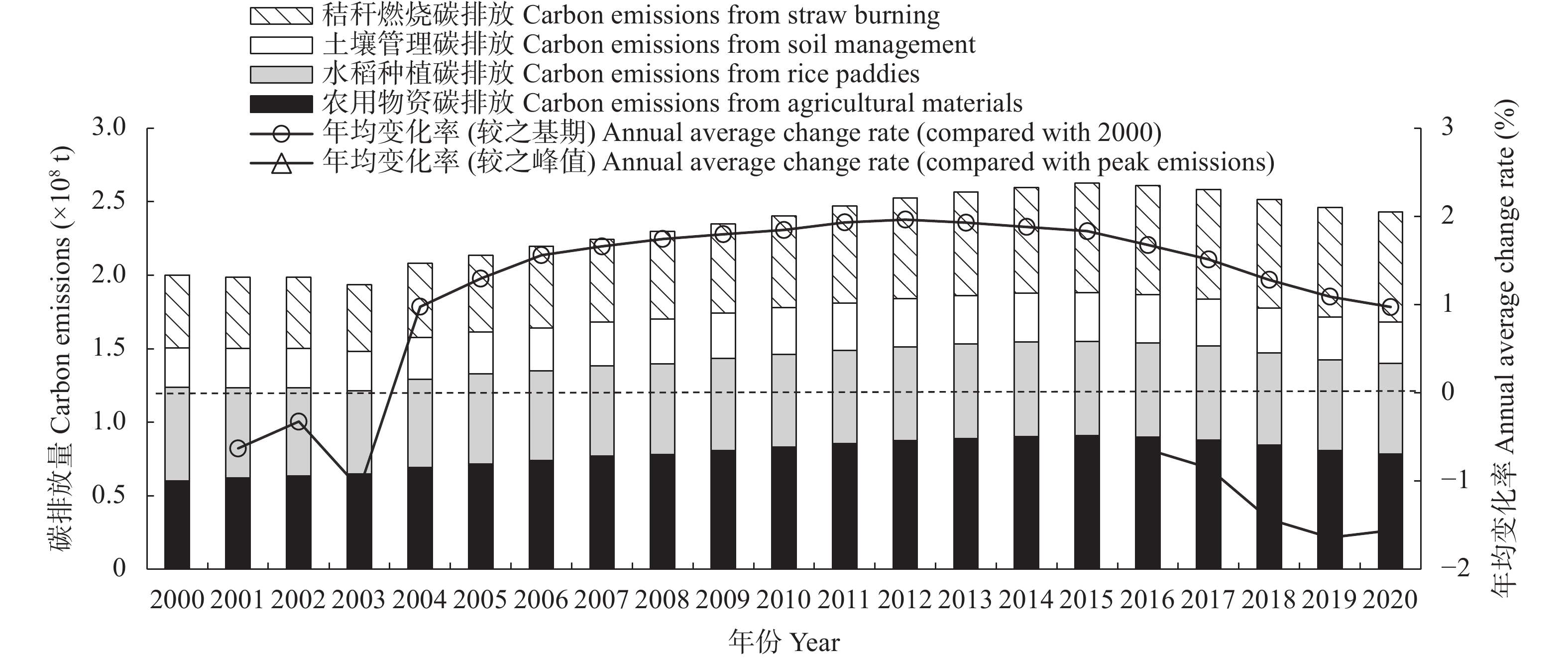
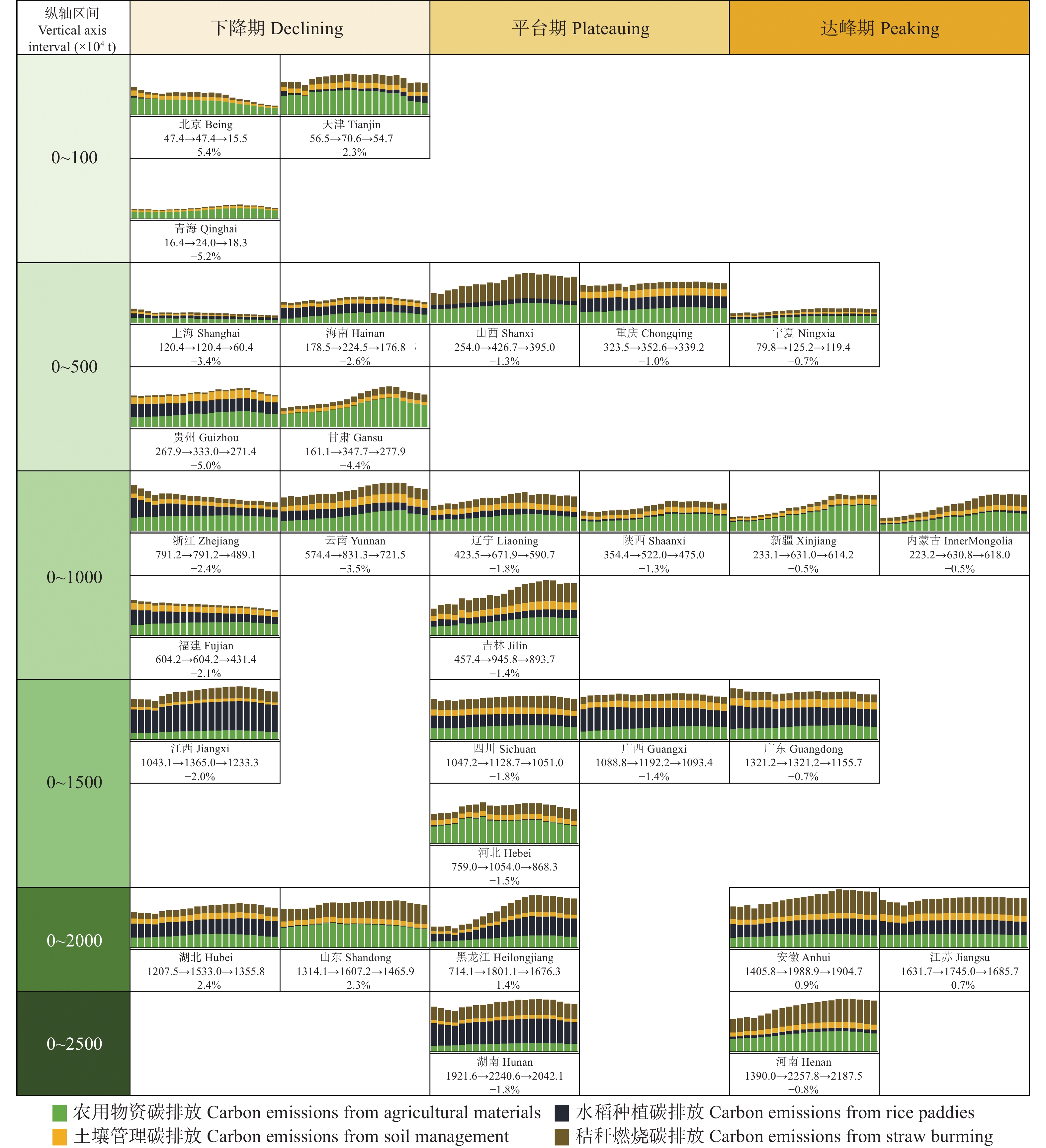
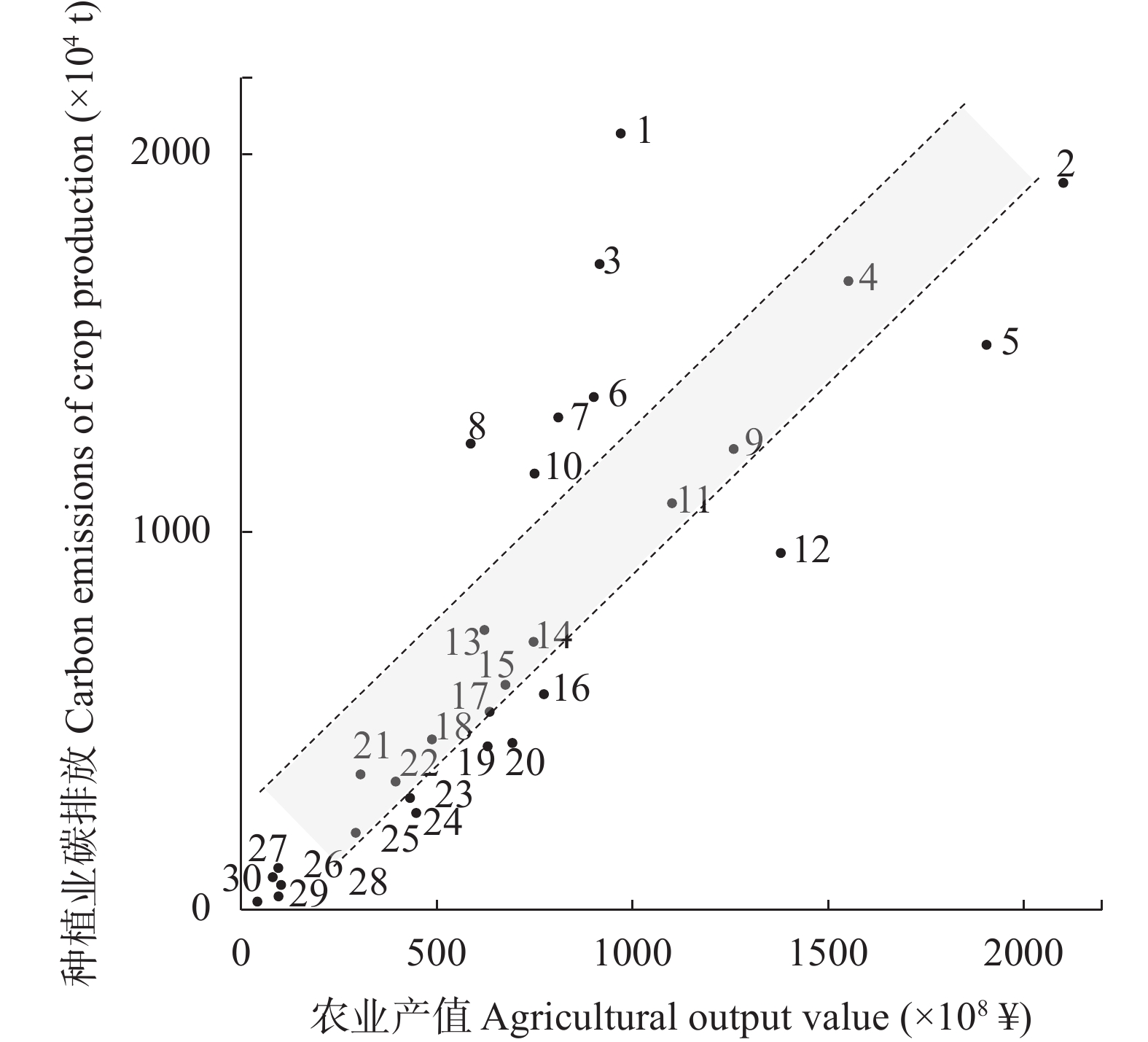
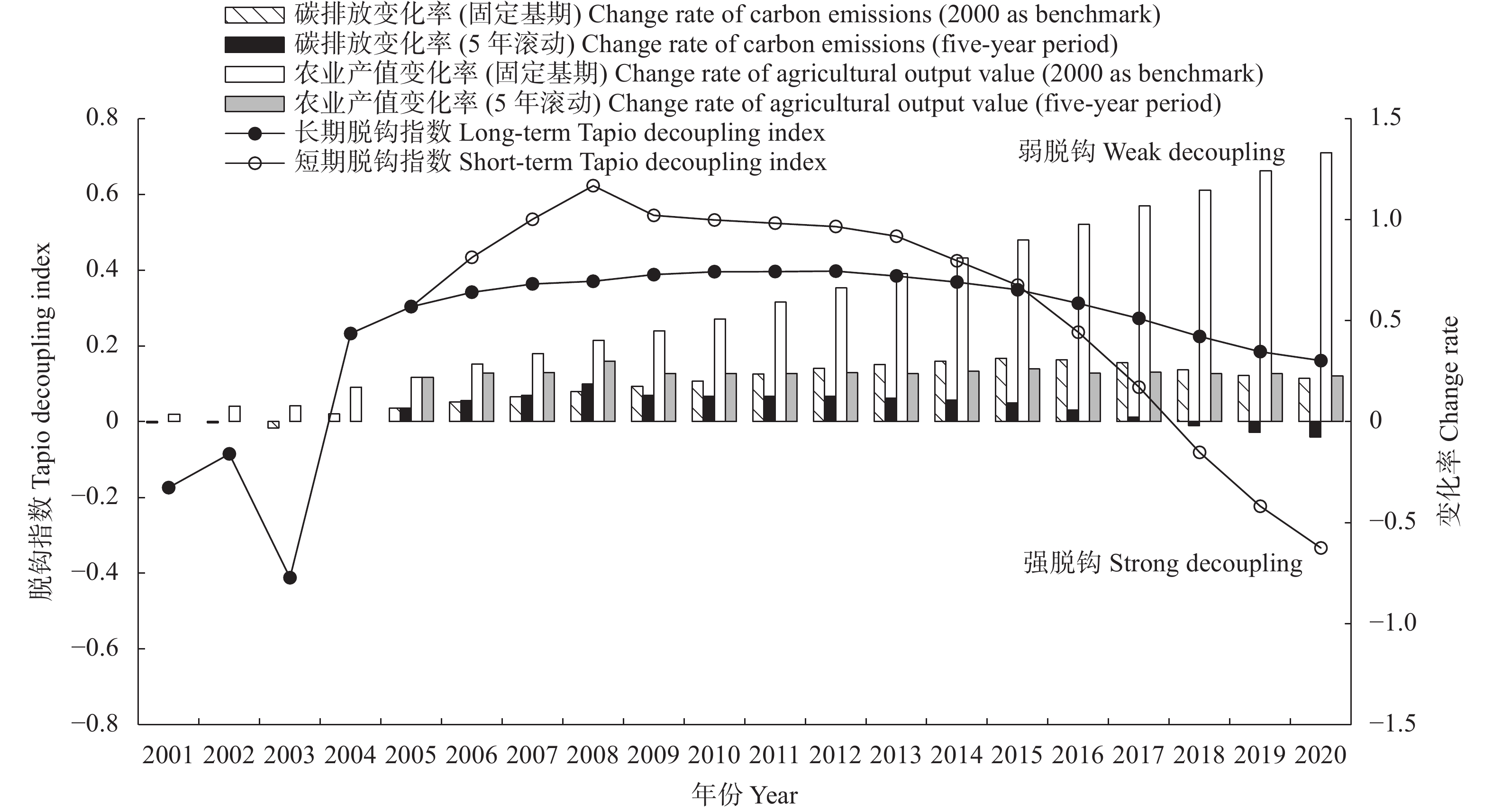
判断种植业碳达峰进程, 可为温室气体减排提供农业领域的数据支撑。考虑农用物资、水稻种植、土壤管理和秸秆燃烧4类排放源, 本文对2000—2020年中国30省(市、自治区)种植业碳排放进行核算, 分类别、分量级对达峰进程展开初步探索, 利用Tapio脱钩指数探讨种植业碳排放与农业产值之间的关系。结果显示: 全国种植业碳排放量年均为23 326.860万t, 在2015年达到峰值26 264.777万t, 达峰后年均变化率为−1.560%, 尚处于平台期。根据达峰进程, 可将30省(市、自治区)分为下降期(北京、天津等13地)、平台期(山西、重庆等10地)、达峰期(河南、安徽等7地)。从全国层面来看, 种植业碳排放与农业产值的长期关系表现为弱脱钩, 短期关系已由弱脱钩转变为强脱钩。就省域层面而言, 短期关系自多种类型并存格局演化为强脱钩主导的极化态势。应根据达峰阶段及特点, 分区域、分类型制定全局减排策略, 加快我国种植业碳排放达峰转降进程。
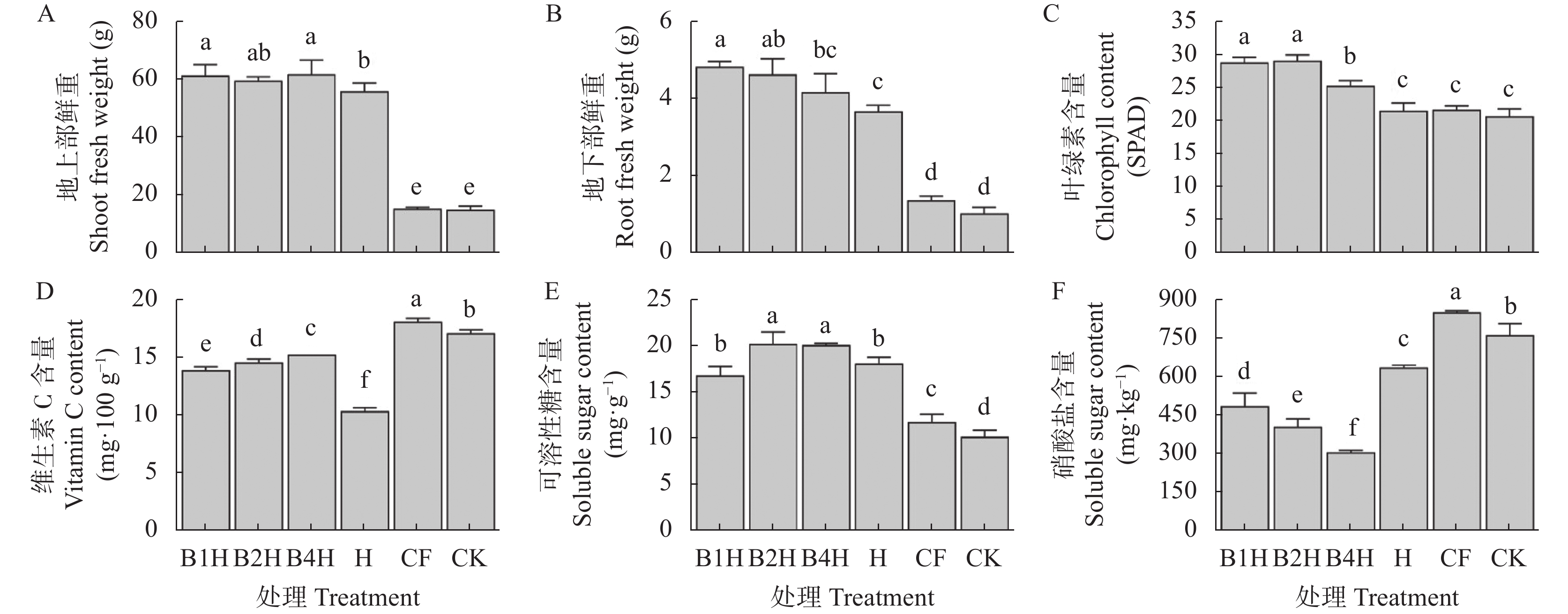
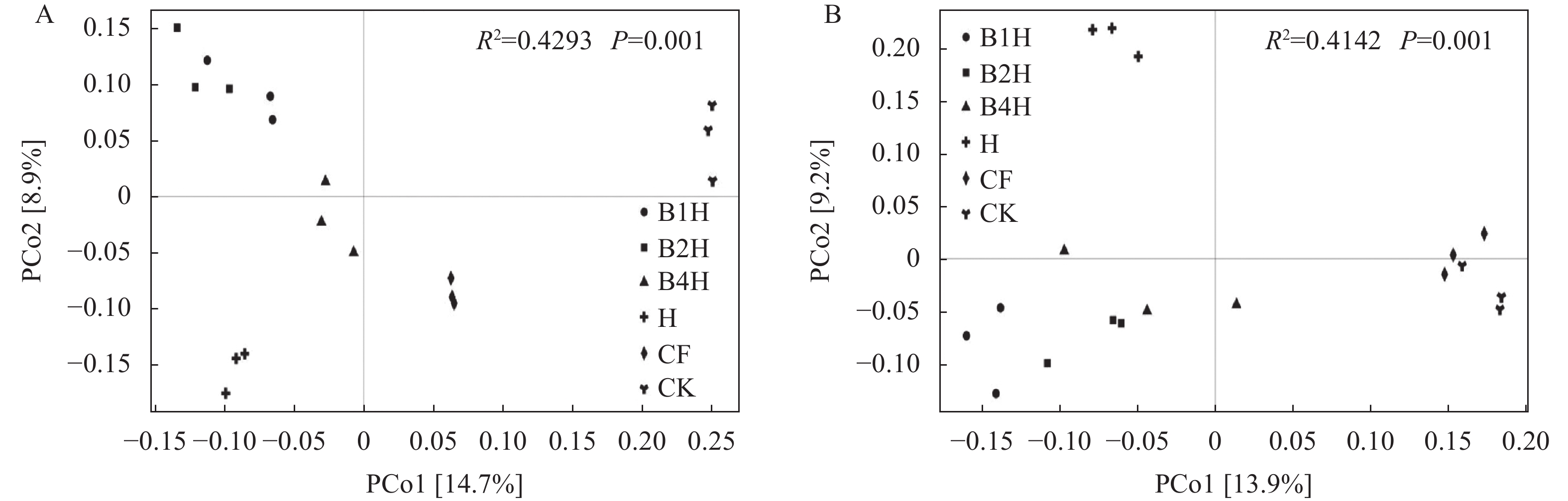
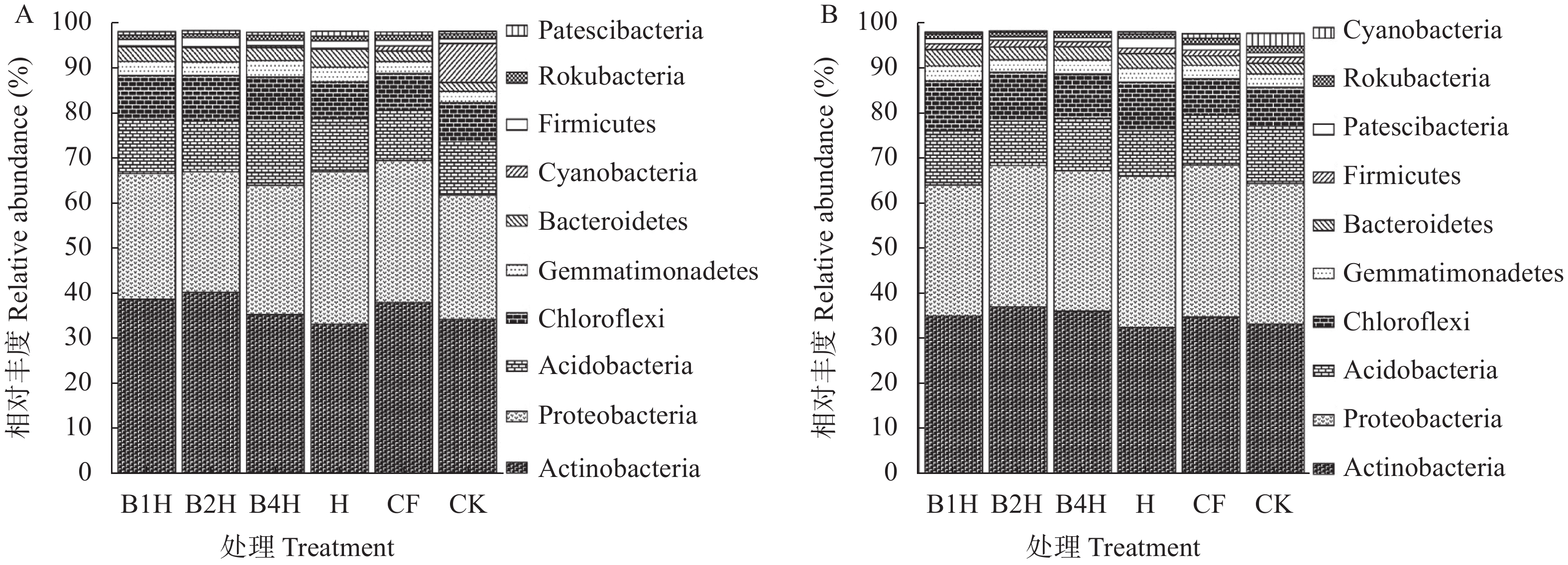
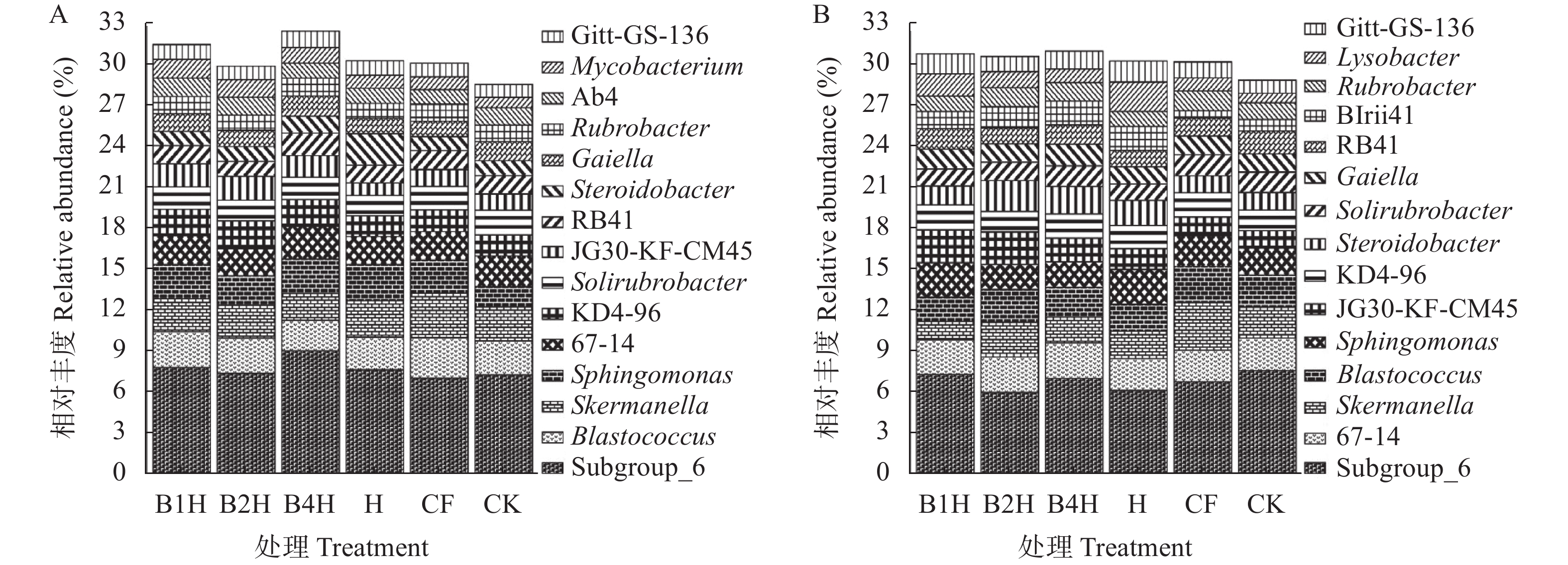
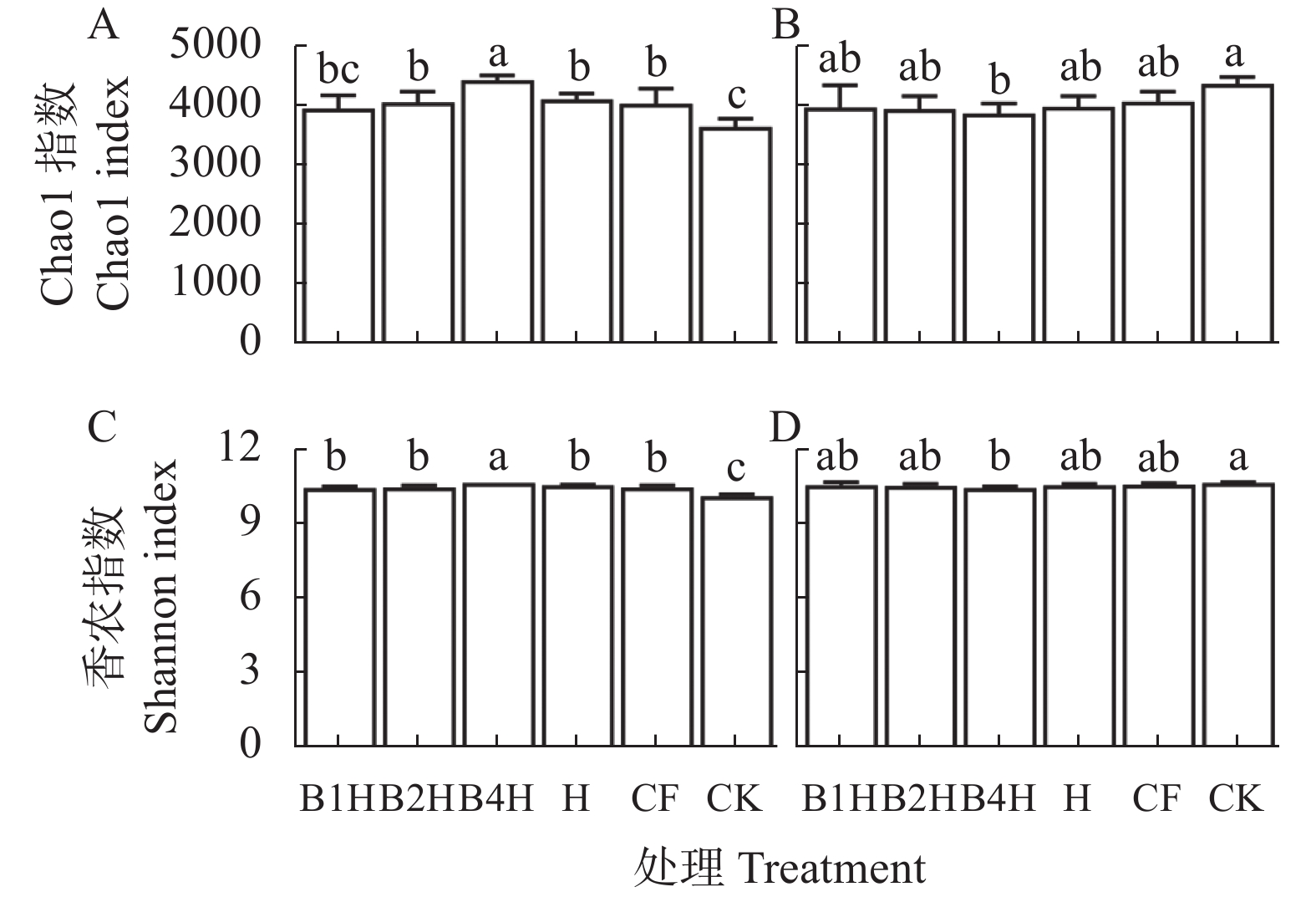
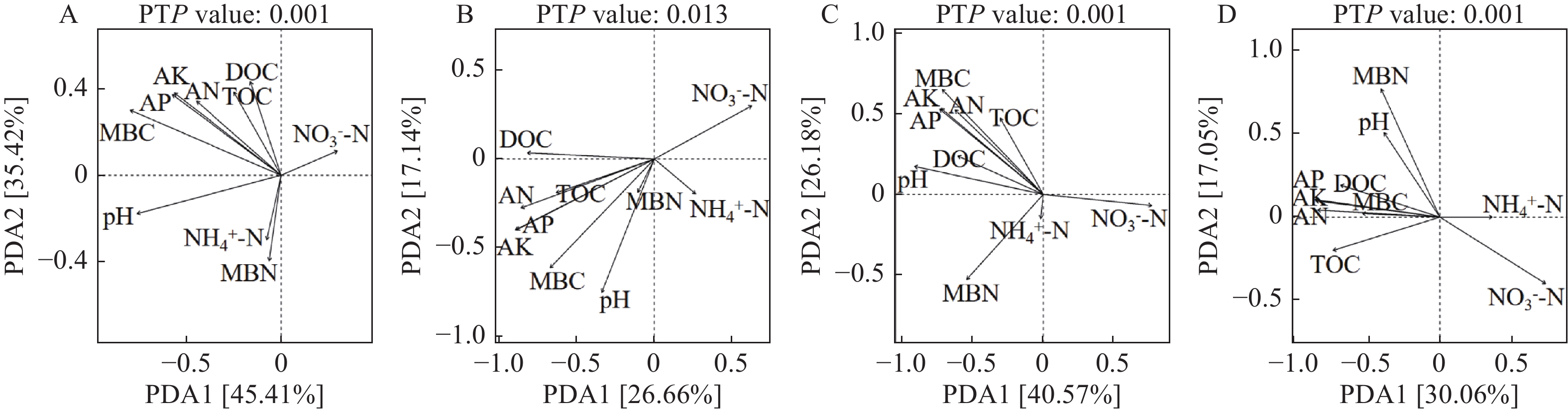
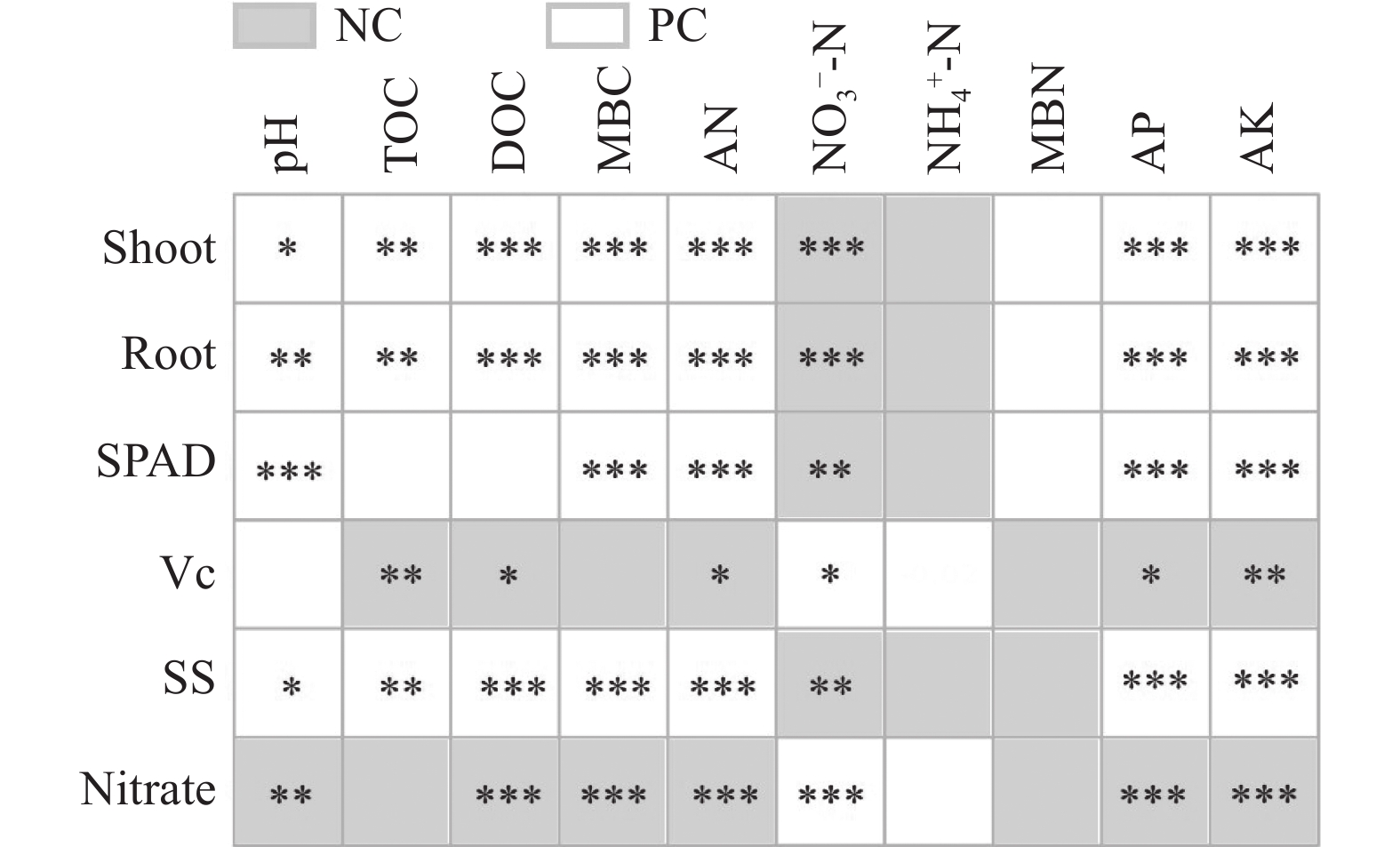
不合理施肥导致土壤退化和产出下降, 废弃物处理则关系到环境和资源可持续发展。生物有机肥在植物促生和土壤培肥方面优势突出, 有利于资源高效利用、减施化肥和蔬菜产业发展。本研究通过向中药渣有机肥分别接种地衣芽孢杆菌、解淀粉芽孢杆菌、巨大芽孢杆菌后发酵制备试验用生物有机肥, 开展生菜温室盆栽试验, 探究不同生物有机肥配施化肥对生菜生长和土壤环境的影响。设置6处理, 分别为3种生物有机肥替代80%化肥氮处理[地衣芽孢杆菌+中药渣有机肥+化肥(B1H)、解淀粉芽孢杆菌+中药渣有机肥+化肥(B2H)、巨大芽孢杆菌+中药渣有机肥+化肥(B4H)]、1种有机肥替代80%化肥氮处理[中药渣有机肥+化肥(H)]和单施化肥处理(CF)以及不施肥处理(CK)。测定和分析收获期生物量和品质等生菜生长指标以及理化性质和细菌多样性等土壤环境指标。结果发现: 各施肥处理对生菜综合增产提质效果以及对土壤养分环境的综合改善效果排序均为B4H>B2H>B1H>H>CF>CK, 与H相比, B4H的生菜地上部鲜重以及叶绿素、维生素C和可溶性糖含量分别提高10.69%、17.77%、47.54%和10.95%, 硝酸盐含量降低52.00%, 土壤有效磷(AP)、速效钾(AK)、碱解氮(AN)、微生物量碳(MBC)和水溶性有机碳(DOC)含量分别提高47.57%、10.98%、8.05%、35.54%和16.10%, 根际土细菌物种丰富度和多样性也分别提高7.68%和0.85%。土壤AP、AK和AN是生菜生长的最重要影响因子, pH、AP和AK则是土壤细菌群落的最关键调控因子, 生物有机肥配施化肥主要通过调控土壤中这些关键因子影响生菜生长和土壤环境。施肥有助于提高生菜根际土细菌Alpha多样性, 较高的根际土细菌Alpha多样性对生菜生长表现出促进作用。总的来说, B4H对生菜生长和土壤环境的改善效果最佳, 可通过大田试验对其促生培肥功效和环境效应作进一步探究。本研究为蔬菜和生物有机肥产业发展提供了新的理论支持, 有利于绿色可持续发展战略实施。



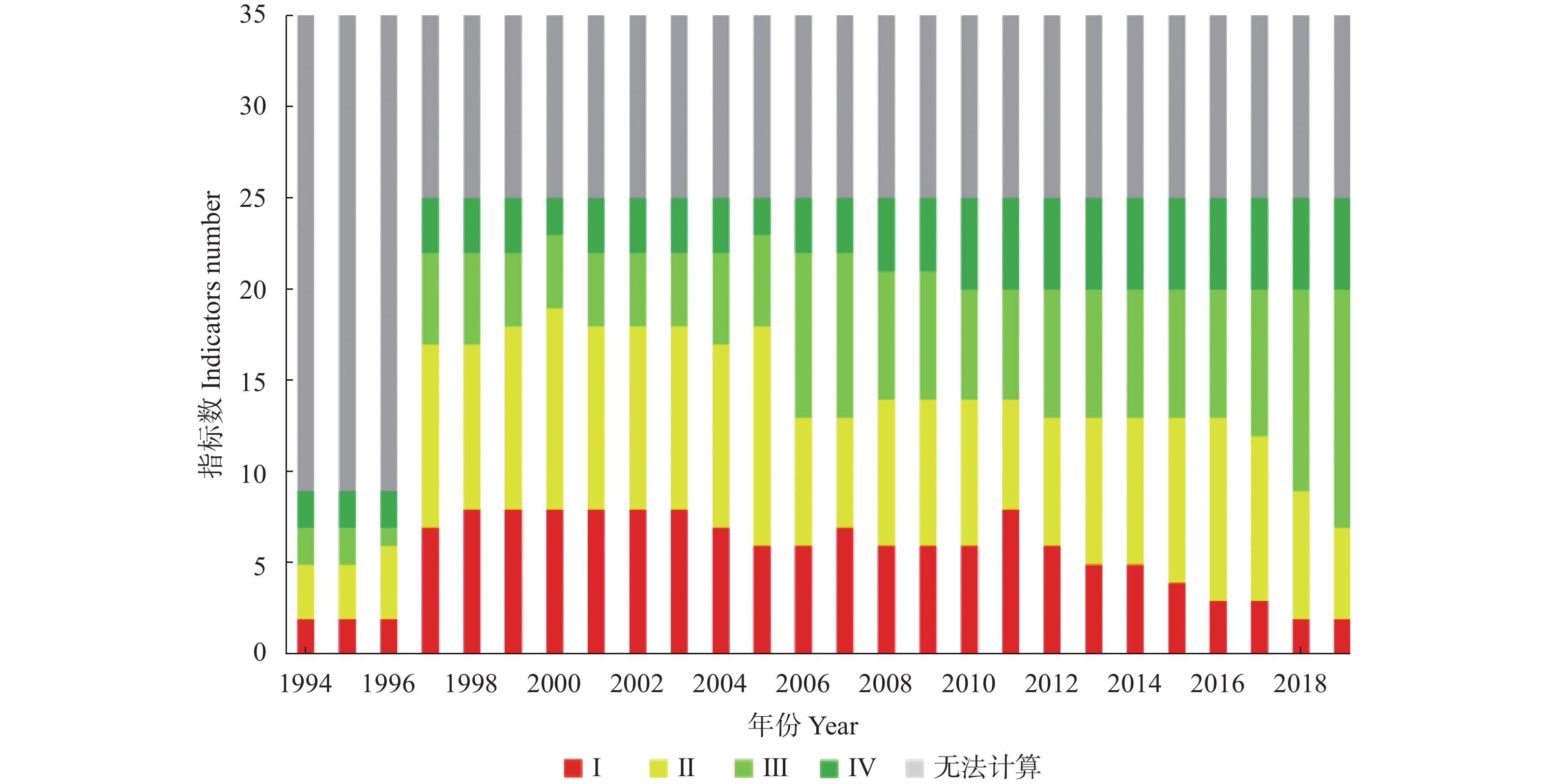
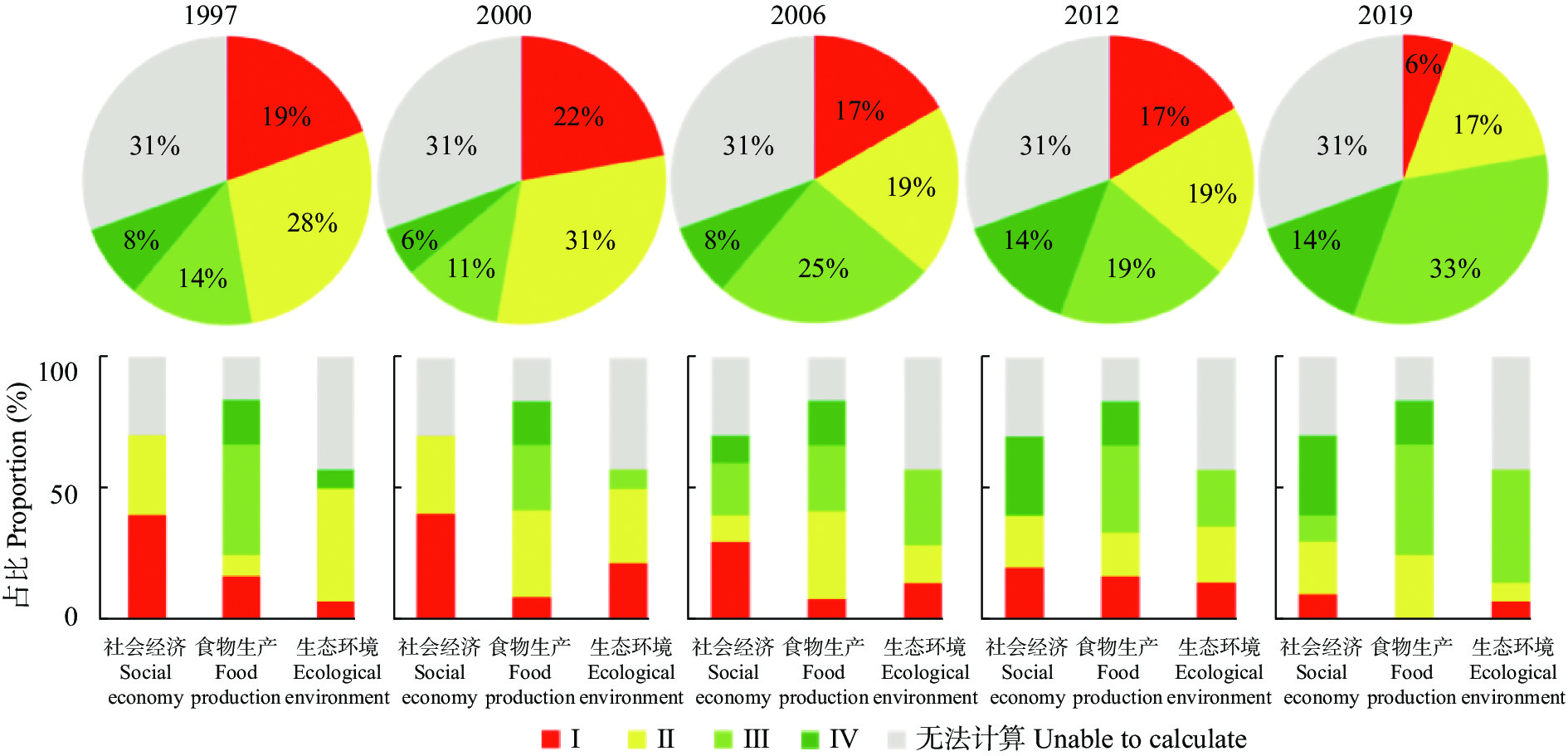
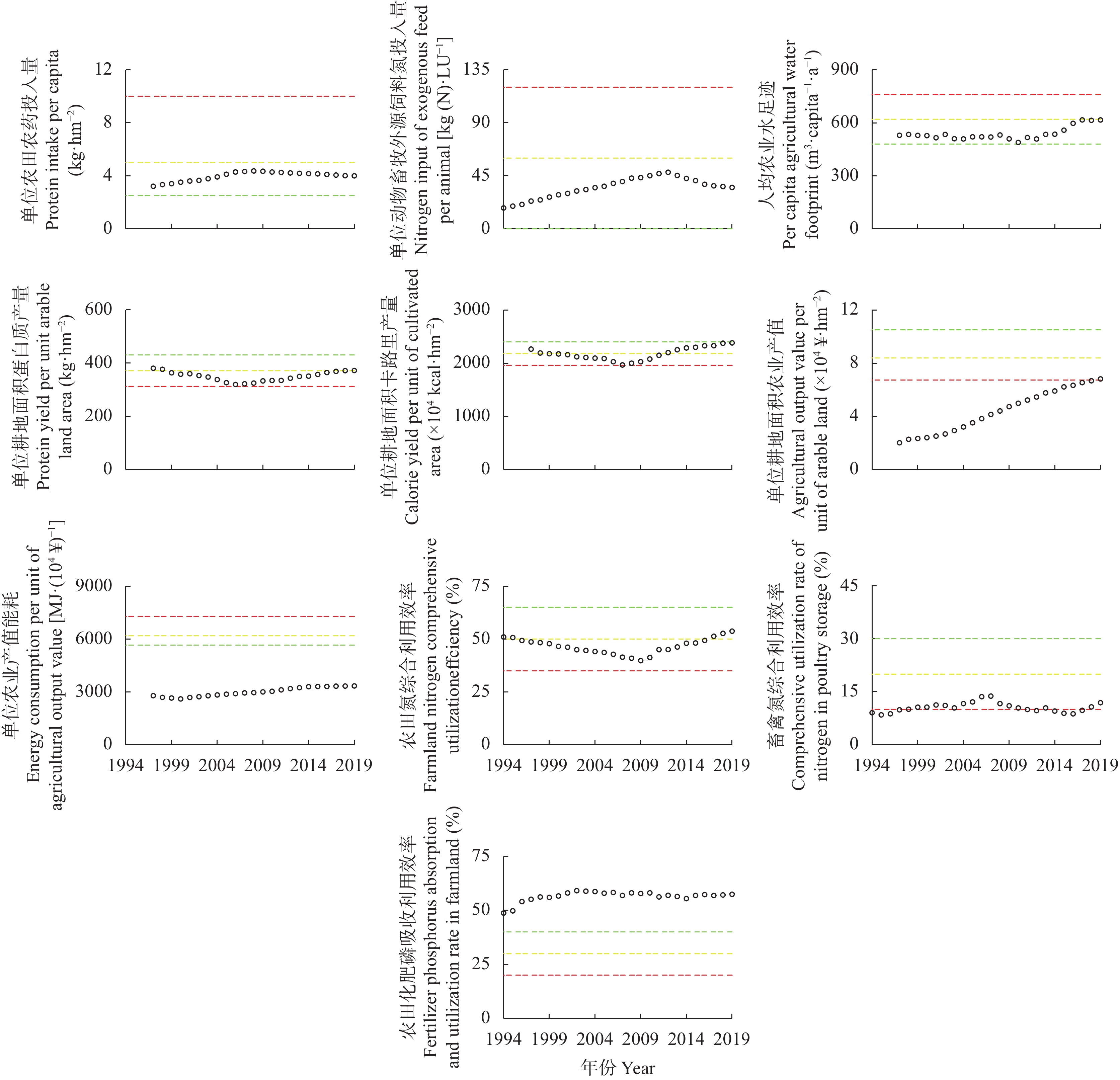
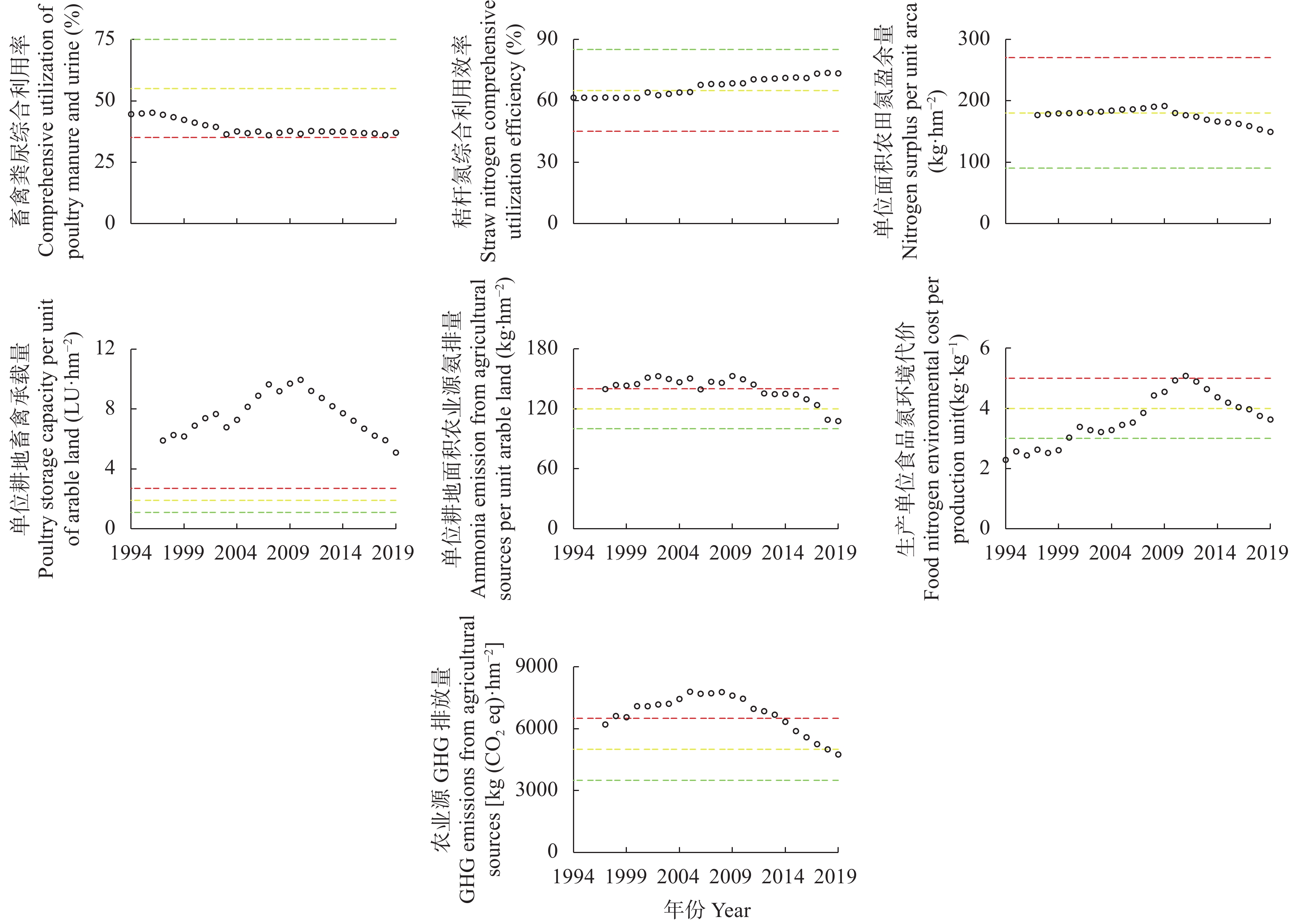
吉林省梨树县作为全国农业生产先进县, 始终走在农业绿色发展的前沿, 本研究将其作为典型案例, 通过已构建的中国农业绿色发展指标体系, 结合NUFER模型, 从社会经济、食物生产、生态环境3个方面, 分析1994—2019年梨树县农业绿色发展指标特征及其变化规律, 进一步探究吉林省县域农业绿色发展的推动及制约因素。结果表明, 1994—2019年梨树县农业绿色发展整体水平稳步提升, Ⅰ、Ⅱ级指标占比由47%下降到23%, Ⅲ、Ⅳ级指标占比由22%上升到47%。在社会经济方面, 人均农业综合投入和农村居民人均可支配收入逐年增加, 2019年较1994年分别增长454.6%和866.7%; 人均蛋白摄入量和动物生产蛋白占比有良好的改善, 均达到Ⅳ级水平; 农业机械化动力虽呈增长趋势, 但仍处在Ⅰ级水平, 有待提高。在食物生产方面, 单位农业产值能耗和农田化肥磷吸收利用效率多年来一直处于Ⅳ级水平, 但畜禽氮综合利用效率始终处在低级水平; 此外, 农药、化肥氮投入均在2019年达到Ⅲ级水平, 但过程中不是持续增长, 受年限的影响较大。在生态环境方面, 截止到2019年, 单位耕地面积因氮投入产生的氮排放、氮盈余及环境代价均呈好转趋势, 但畜禽粪尿综合利用率和单位耕地畜禽承载量始终处在Ⅰ级水平, 说明梨树县畜牧业发展仍有很大的挑战。综上, 资源未充分利用、环境污染和生态破坏严重制约吉林省县域农业绿色发展, 究其原因是种植结构单一、优质农产品产量低, 农药、化肥等投入量较高, 畜牧业养殖数量不稳定, 下一步急需合理开发利用黑土地资源, 大力推广测土配方施肥、保护性耕作等优秀农业技术, 解决优质农产品供需矛盾等问题, 减少对生态环境的伤害, 全面推进吉林省县域的农业绿色发展。

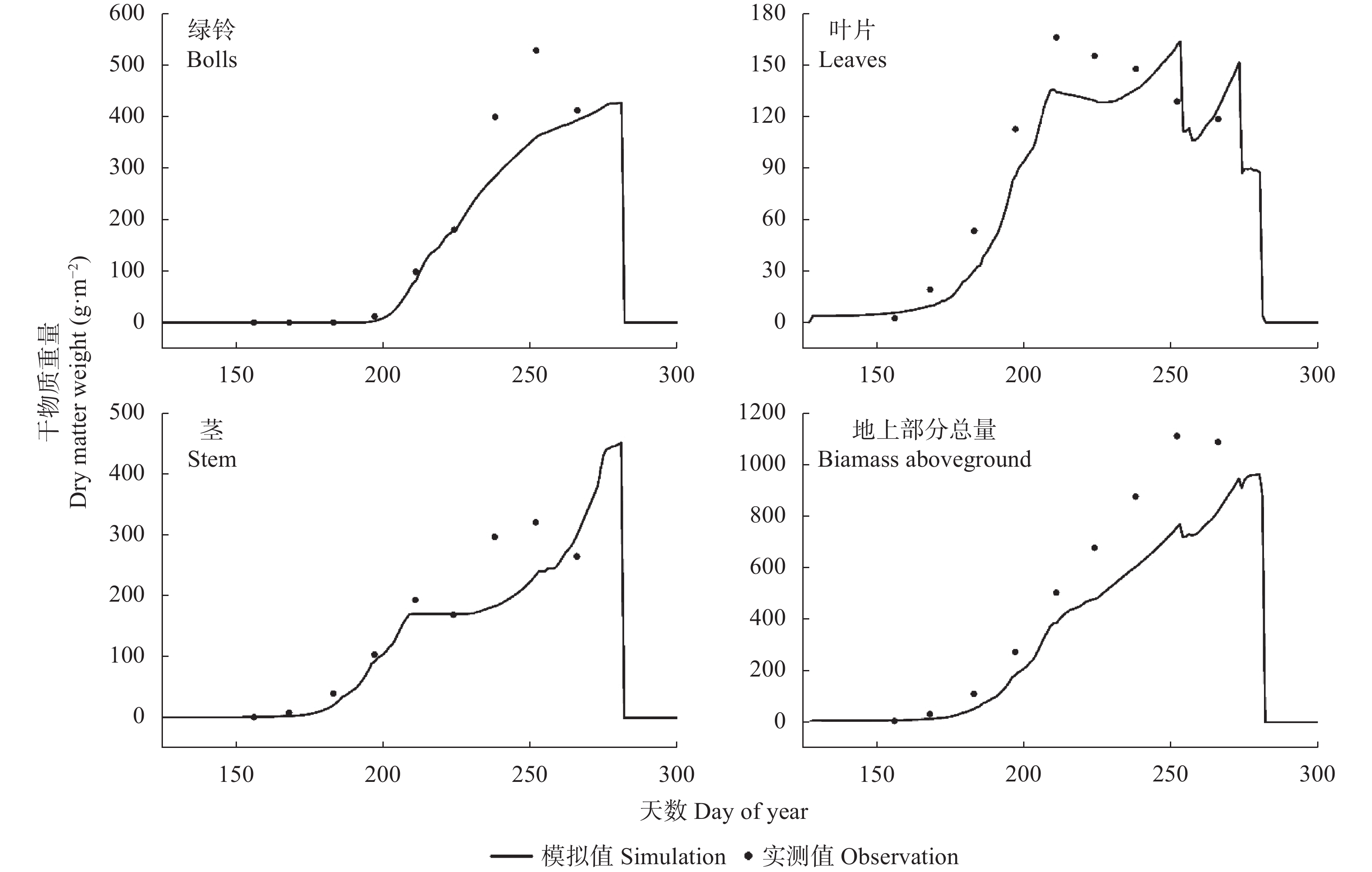
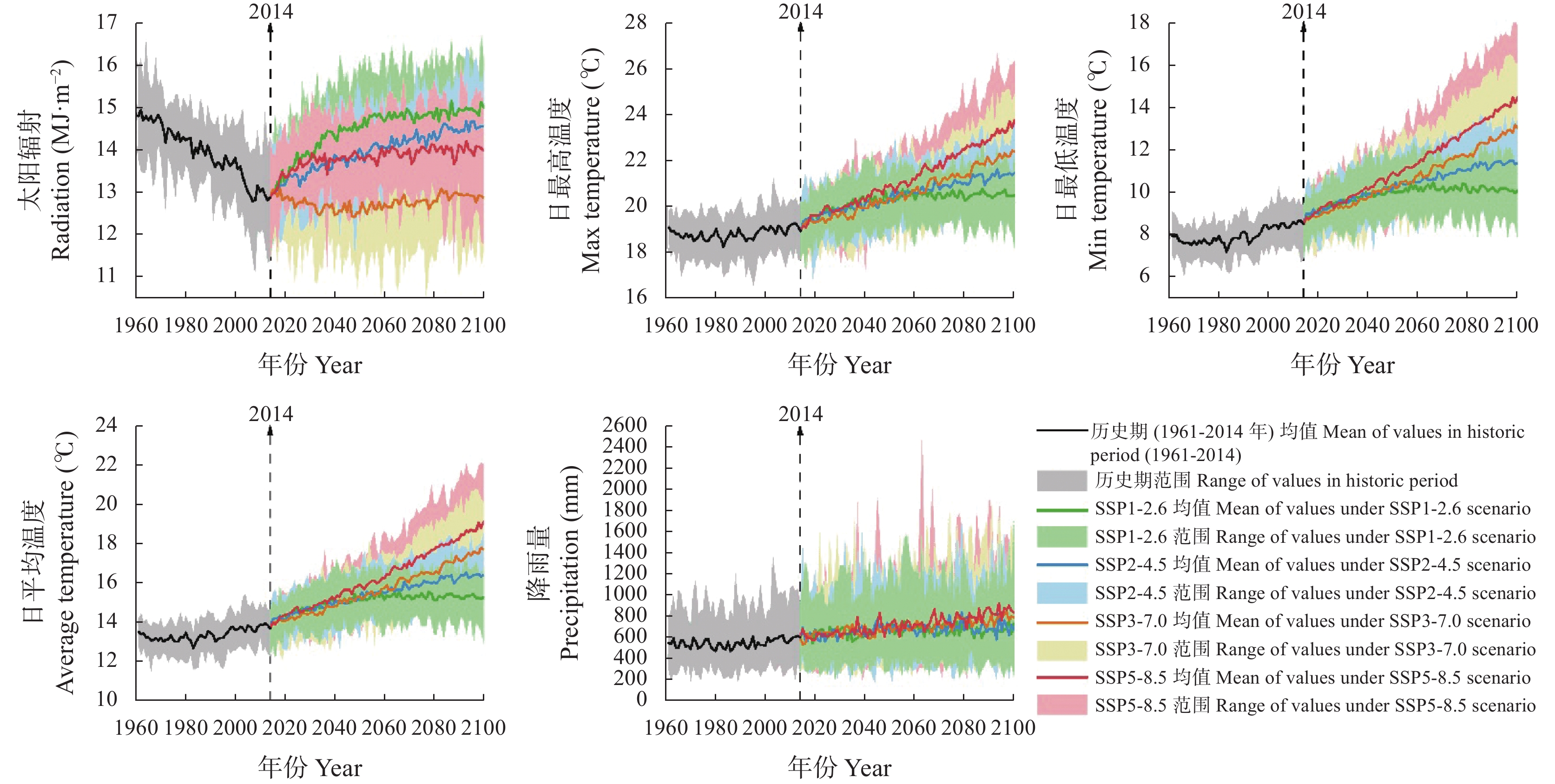
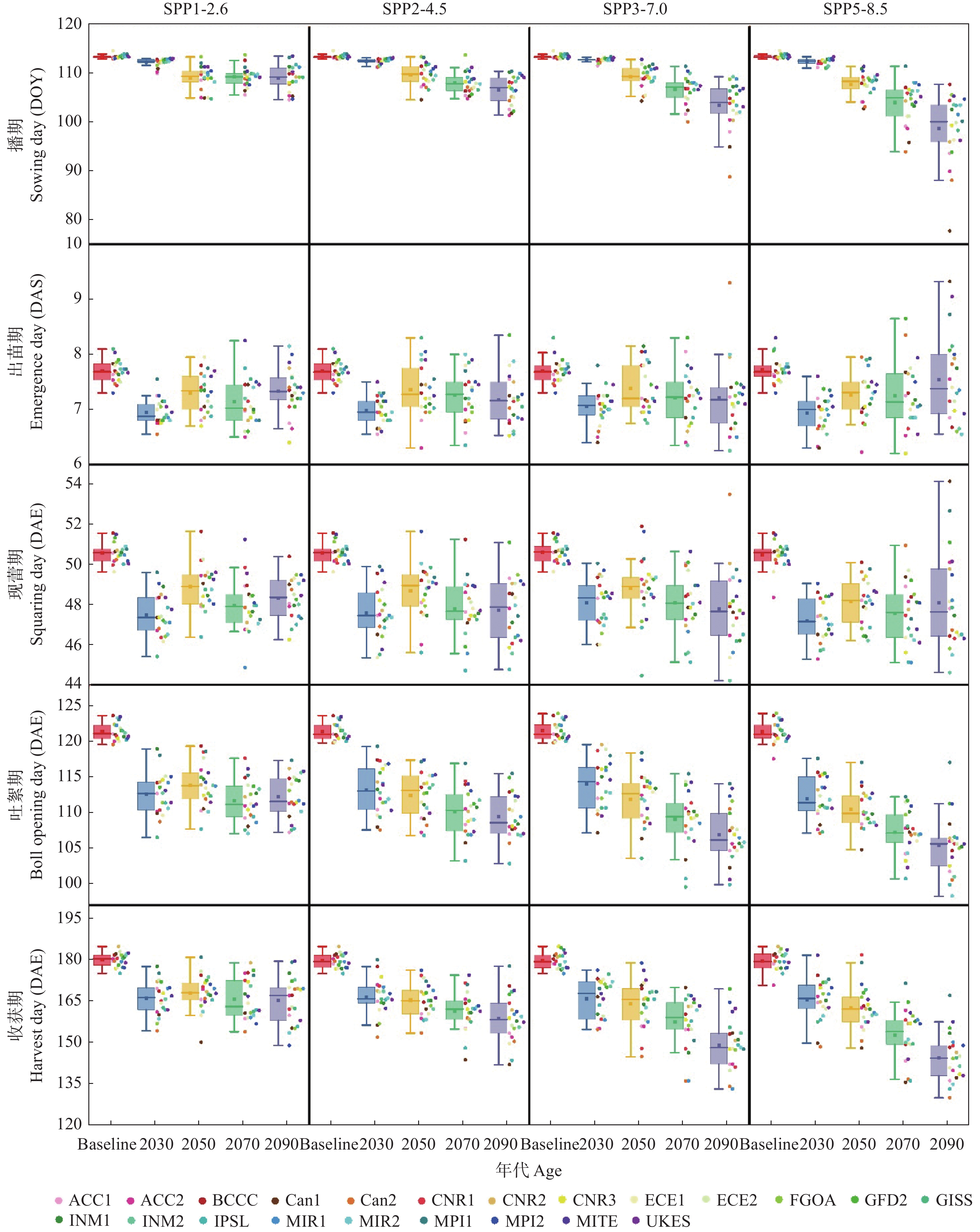
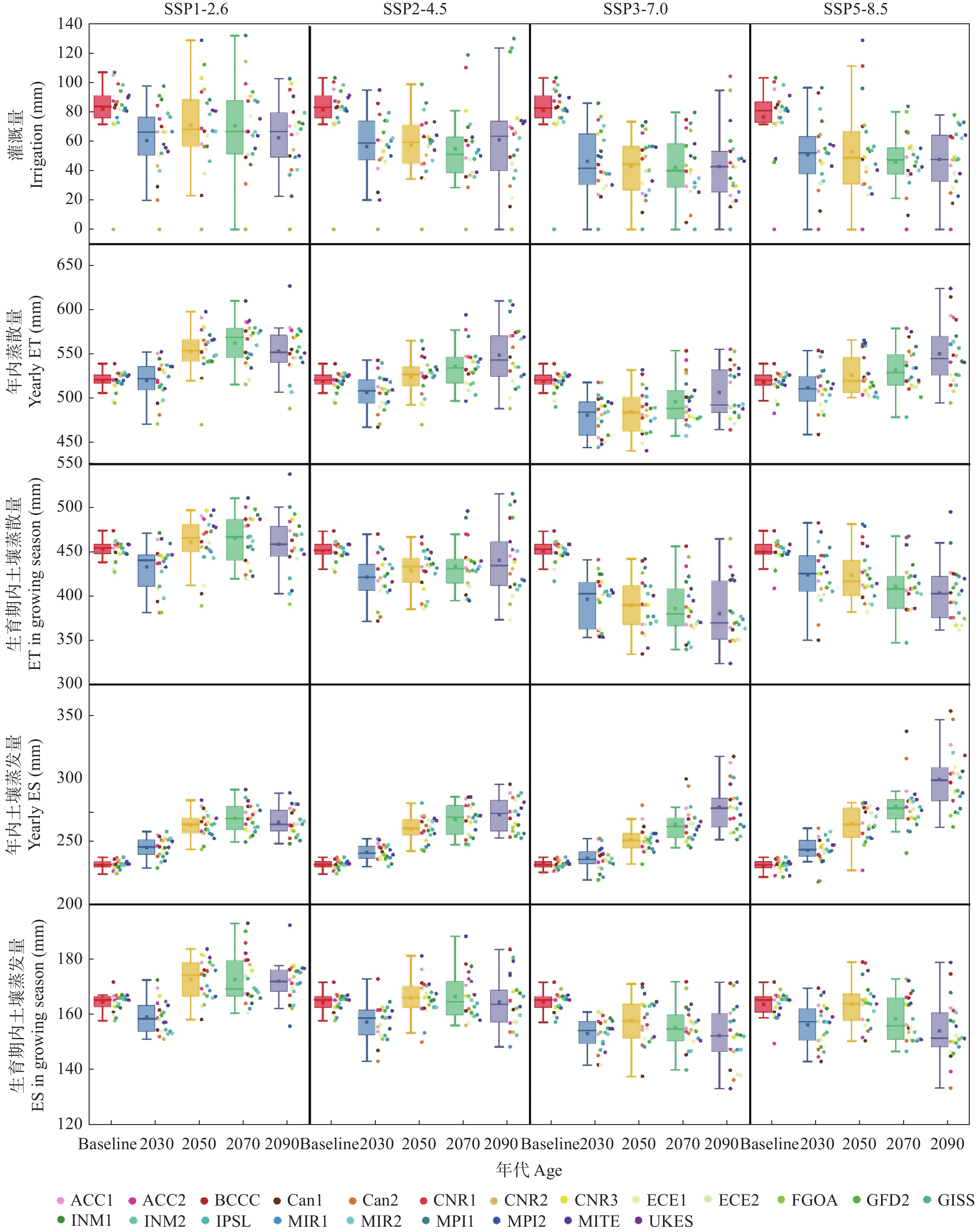
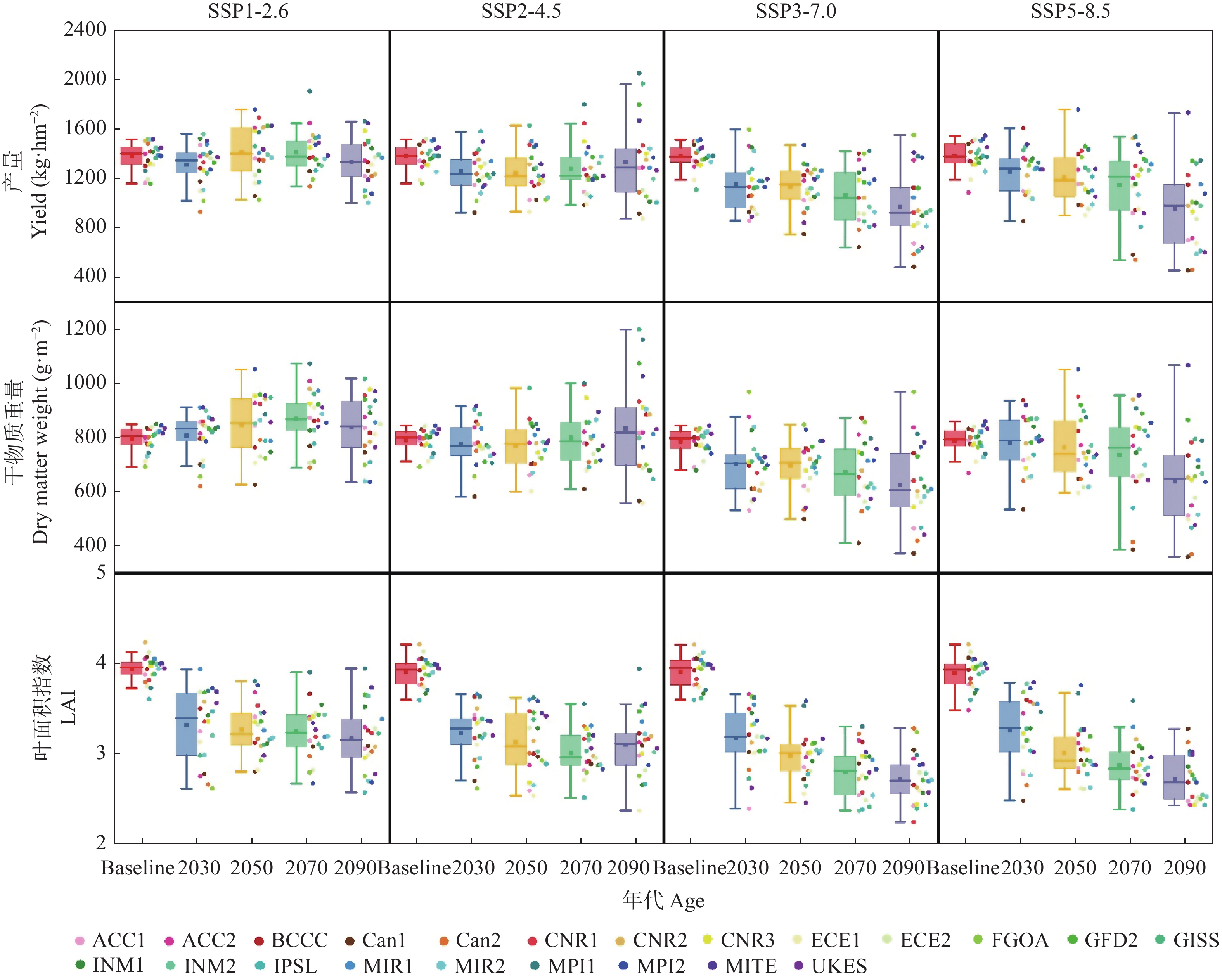
气候模式是气候变化影响评估中不确定性的主要来源, 前人的研究多采用单个或较少的气候模式进行评估, 采用多种气候模式进行驱动可以降低由于气候模式的选择带来的误差。本研究在两年大田试验的基础上对作物模型APSIM-COTTON进行了精细的校验, 并选择22个GCMs(Global Climate Models)模式驱动作物模型评估了气候变化对河北棉花生产和耗水的影响。结果显示, 在所有气候情景下, 未来所有时间段, 播期提前, 各个发育时期(出苗、现蕾、吐絮、成熟)都较基准期缩短, 例如收获期在2090s 年代的SSP1-2.6、SSP2-4.5、SSP3-7.0和SSP5-8.5情景下分别提前15.3d、21.0 d、30.3 d和35.2 d。年内总蒸散量在多数情景下呈总的增加的趋势, 在SSP5-8.5情景下2030s、2050s、2070s和2090s分别增加6.5 mm、7.8 mm、14.3 mm和32.7 mm , 而灌水量减少25.7 mm、23.8 mm、30.5 mm和29.0 mm。棉花产量在未来则表现出在低辐射强迫下不同年代差异不大, 而在高辐射胁迫强迫下随着年代增加而降低的趋势。在SSP1-2.6、SSP2-4.5、SSP3-7.0和SSP5-8.5情景下2090s皮棉产量相比基准期分别减少61.5 kg∙hm−2、46.6 kg∙hm−2、407.1 kg∙hm−2和432.5 kg∙hm−2。棉花生产和耗水对未来气候变化的响应是气候要素CO2浓度、太阳辐射强度、温度、降雨等综合作用的结果, 本研究模拟结果为未来农业措施的响应提供理论支撑。

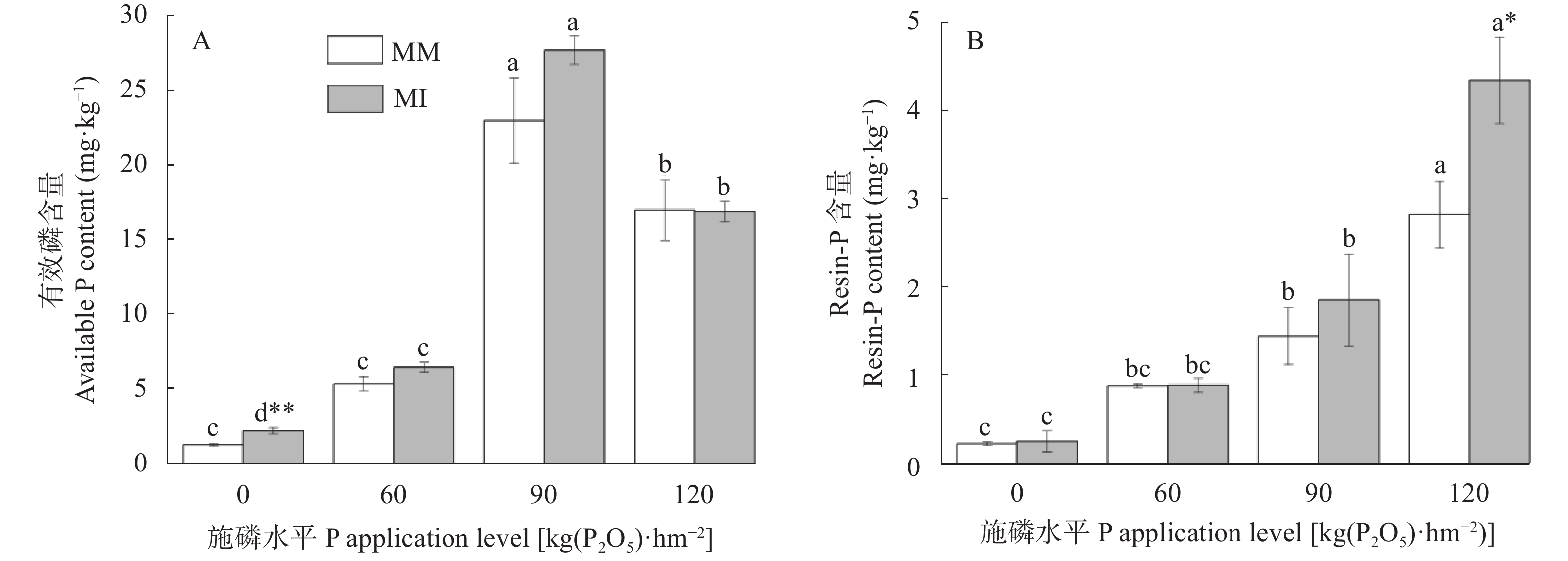
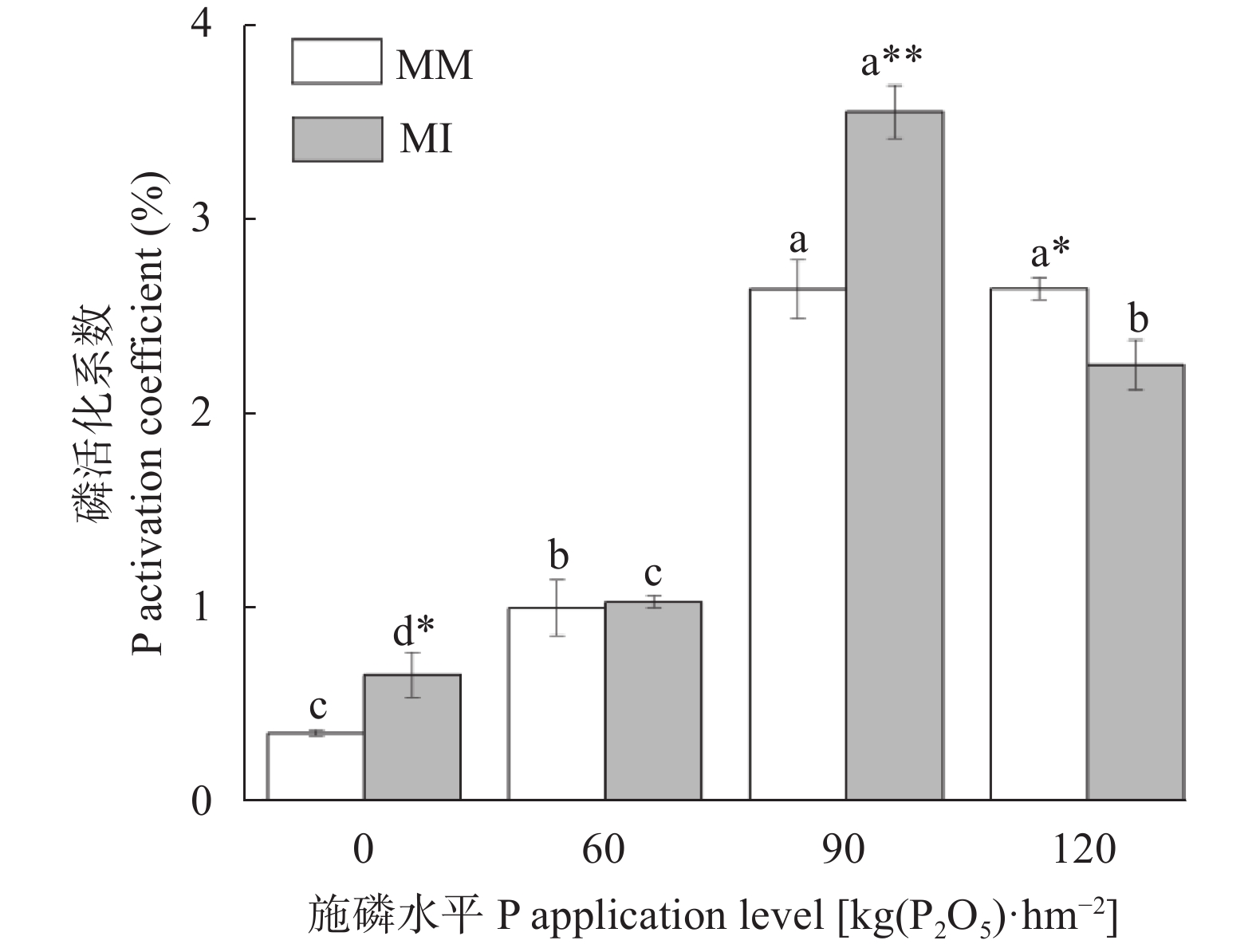
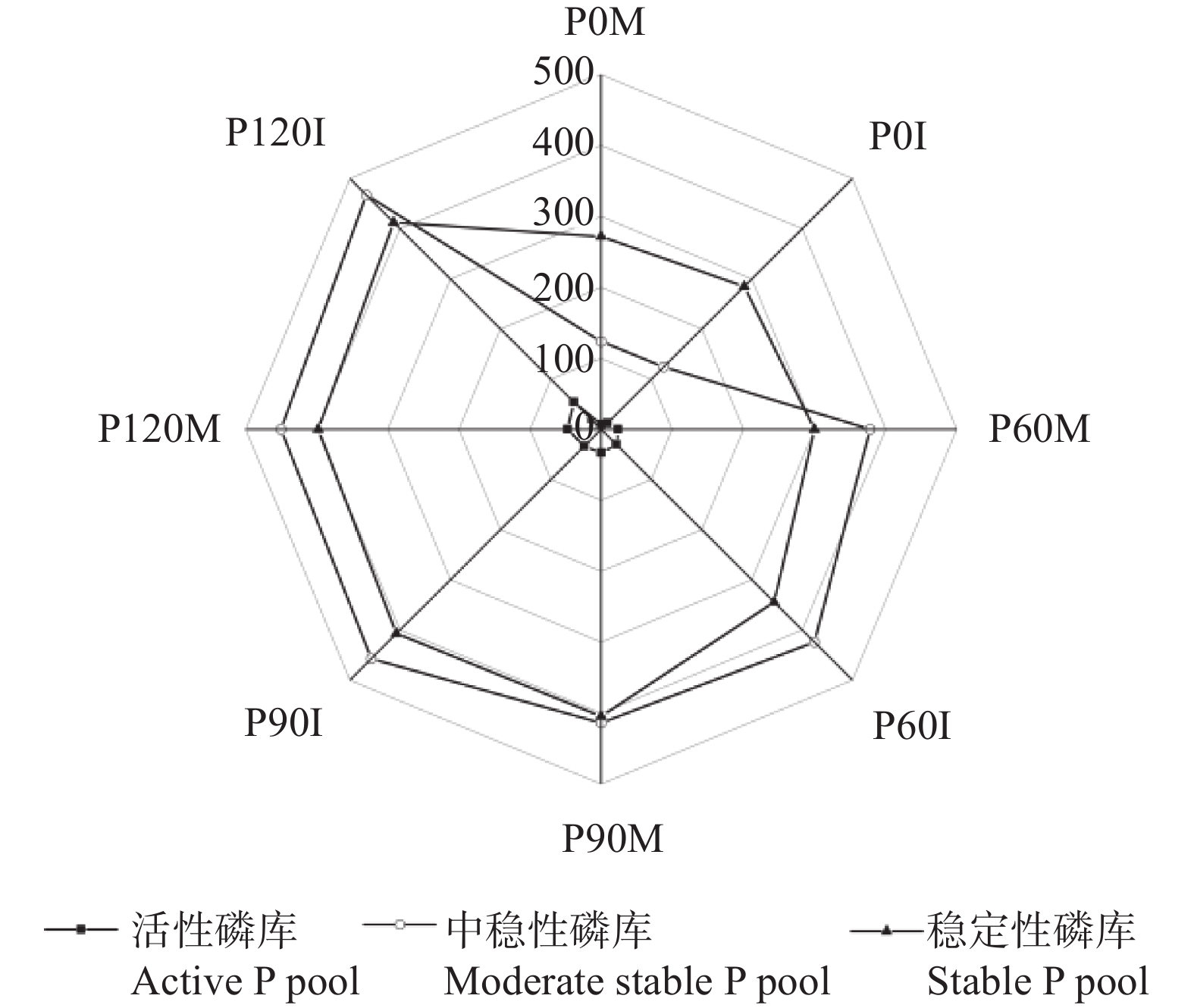
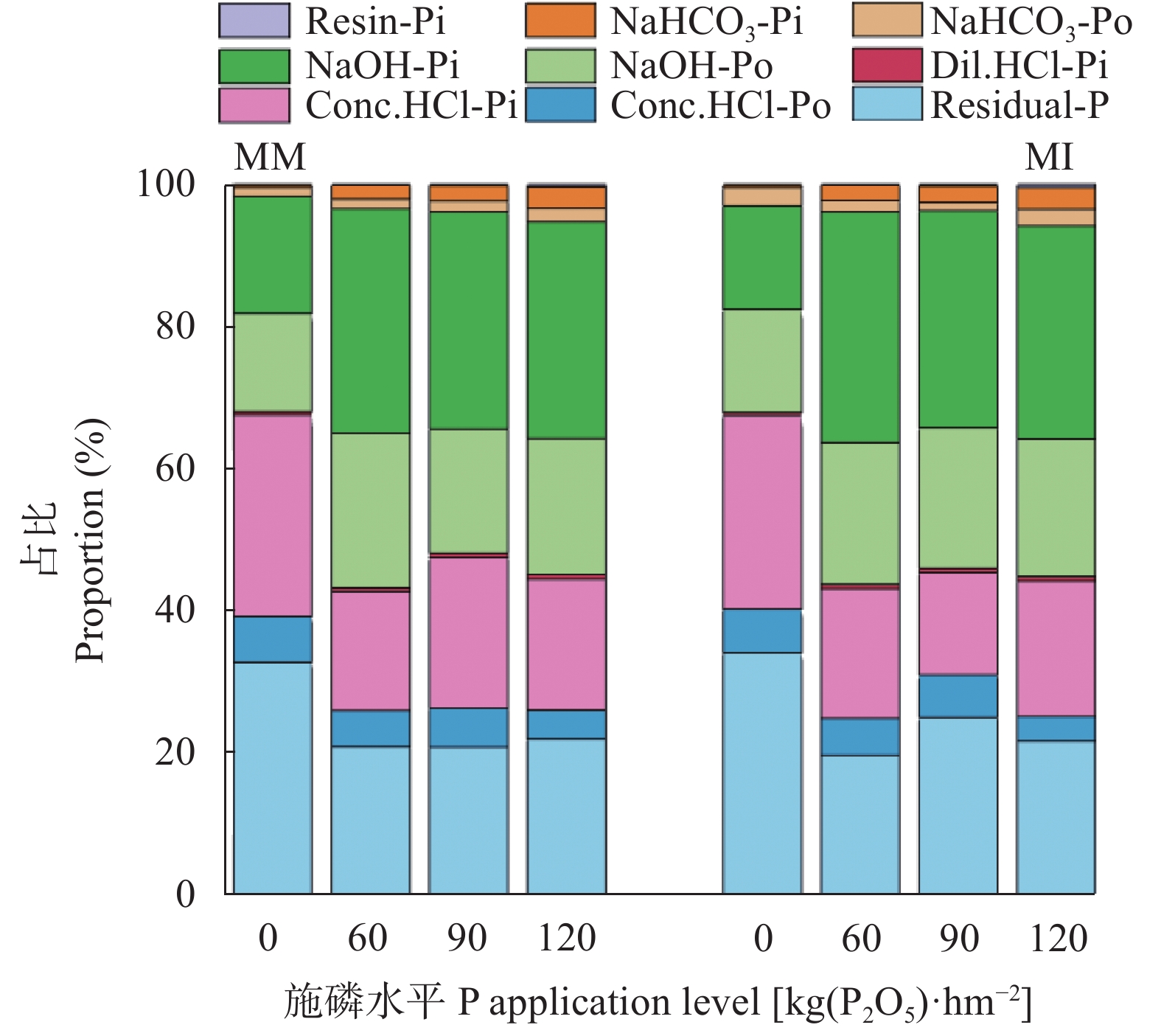
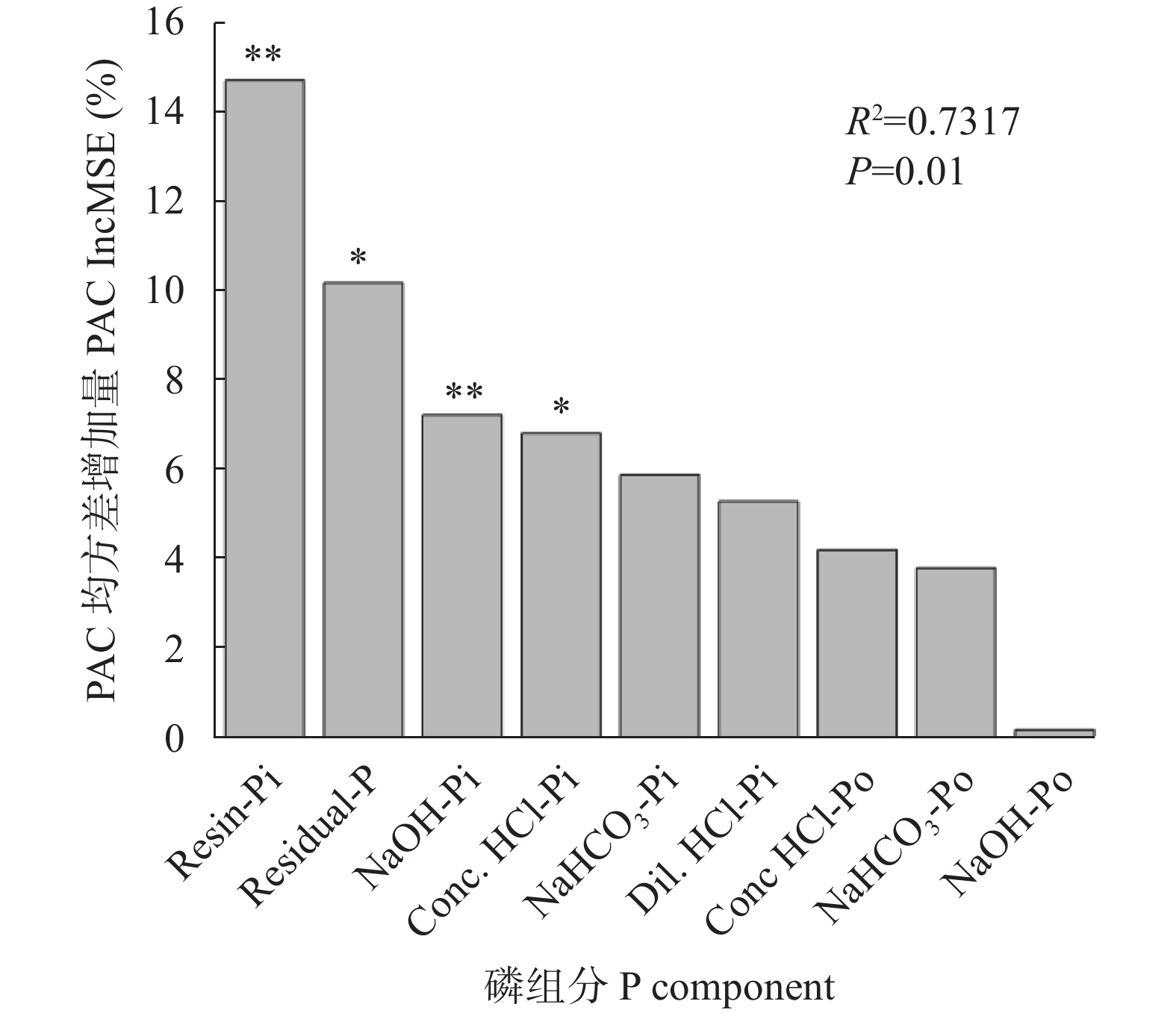
红壤固磷能力强, 但合理间作可促进磷吸收, 减少磷固定。本研究基于连续4年的田间定位试验, 分别设置玉米大豆间作(MI)和玉米单作(MM) 2种种植模式, 不施磷肥(P0)、施P2O5 60 kg∙hm−2 (P60)、施P2O5 90 kg∙hm−2 (P90)及施P2O5 120 kg∙hm−2 (P120) 4个施磷水平, 采用改良的Hedley磷分级法, 研究了玉米大豆间作对玉米根际土壤磷组分的影响及其磷梯度响应; 通过随机森林模型, 探究了不同磷组分对土壤磷活化系数(PAC)的贡献。玉米大豆间作提高了红壤施磷处理的总磷含量和磷有效性。与玉米单作相比, P0水平下间作玉米根际土壤速效磷含量显著提高70.4% (P<0.01)。玉米大豆间作显著促进了红壤磷的活化和向活性磷库的转化。在P0和P90水平下, 间作土壤PAC较单作分别显著提高87.4% (P<0.05)和34.6% (P<0.01)。间作使红壤活性磷库占总磷比例平均提高15.1%。其中无机活性磷组分中Resin-P在P120水平下含量较单作显著提高53.7% (P<0.05), 有机活性磷库中碳 酸 氢 钠 浸 提 有机磷(NaHCO3-Po)含量在P0、P120水平下分别显著提高117.0%、25.6% (P<0.05)。间作使红壤稳定性磷库占总磷比例降低1.1%, 差异不显著。在P90水平下, 稳定性磷库中稀盐酸浸提无机磷(Conc. HCl-Pi)含量较单作显著降低40.2% (P<0.01)。随机森林模型显示土壤无机磷是PAC的主要决定因素, 其中去除水溶性无机磷(Resin-Pi)的预测值时, 土壤PAC的均方差增加14.7%。玉米大豆间作显著提高了玉米根际土壤有效磷含量及土壤PAC, 提高了玉米根际土壤活性磷库、中稳性磷库的比例, 同时降低了稳定性磷库的比例, 玉米大豆间作对磷库的活化在中低施磷水平下作用显著, 在高施磷水平下活化作用不明显, 而其中土壤无机磷组分对PAC影响较大。说明玉米大豆间作促进了红壤磷的活化和向活性磷库的转化, 特别是在中低施磷条件下, 而在高施磷(P120)条件下间作红壤磷的活化作用不明显。
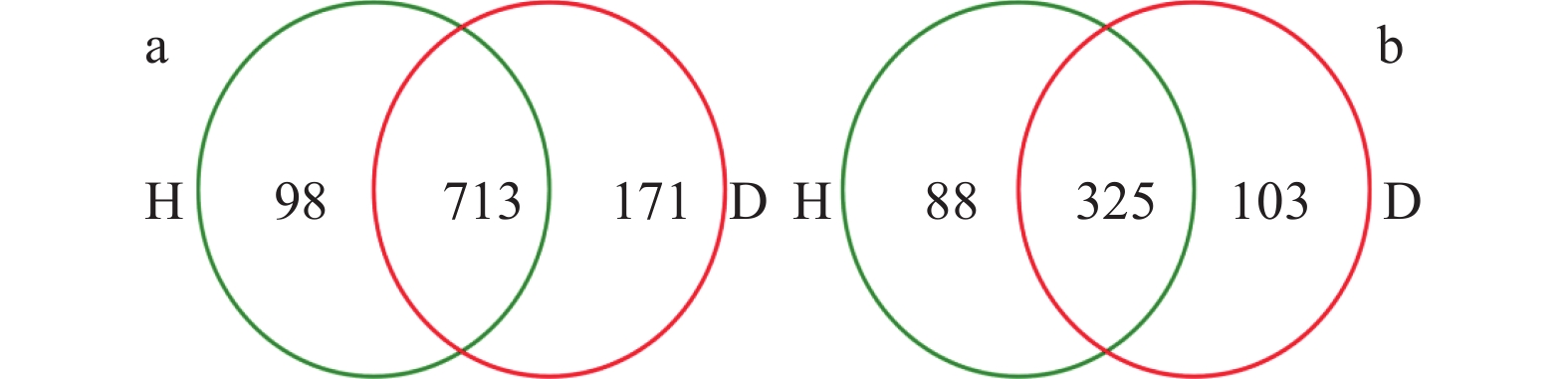
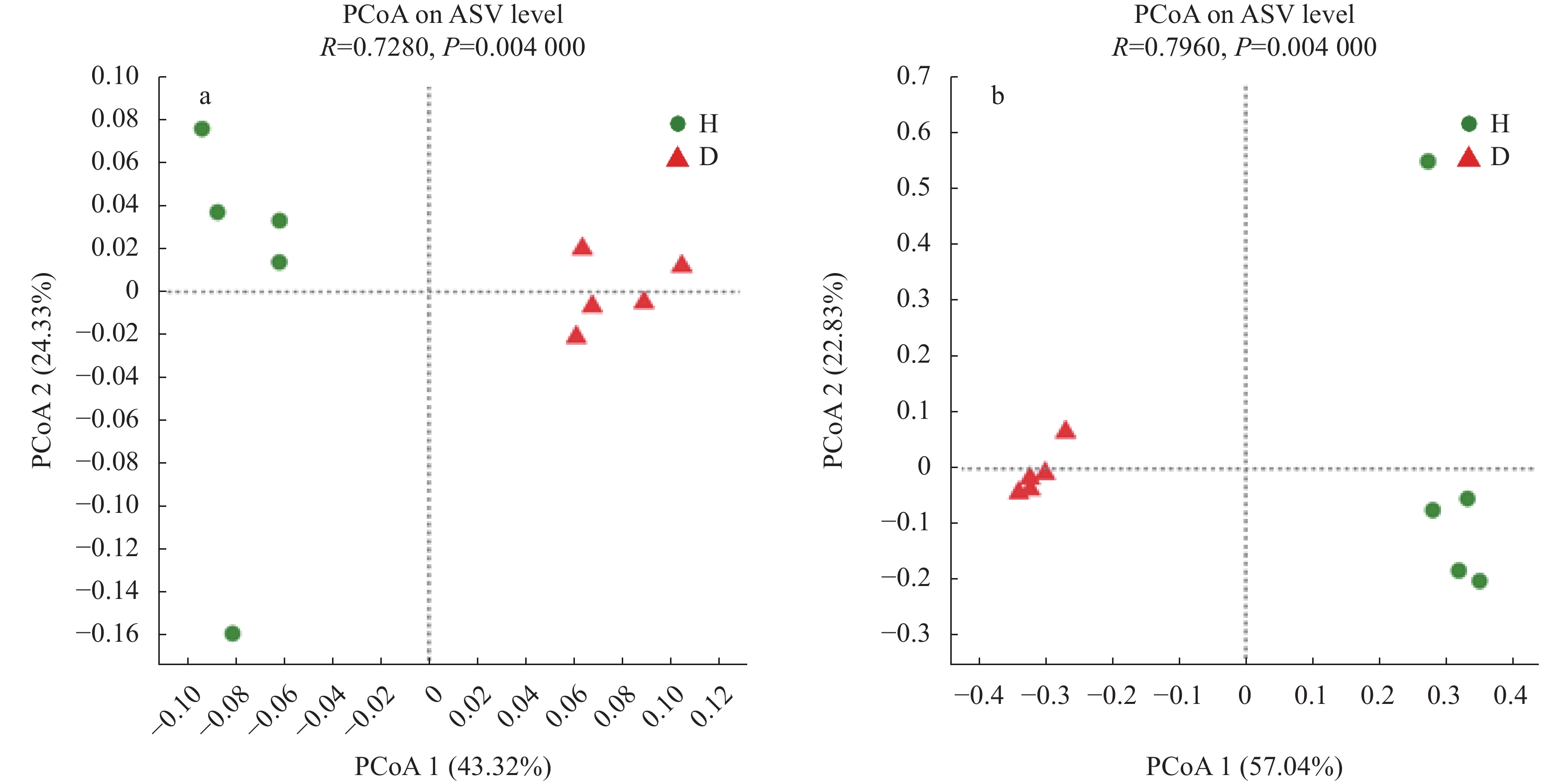
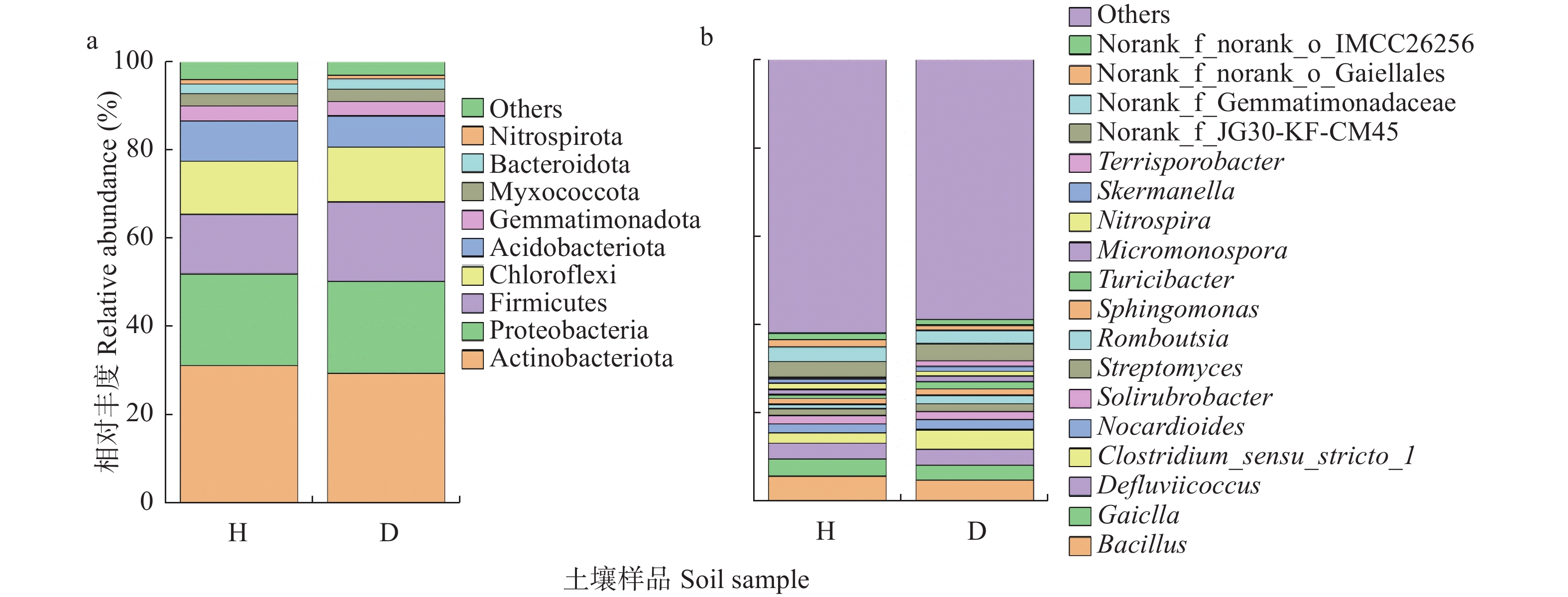
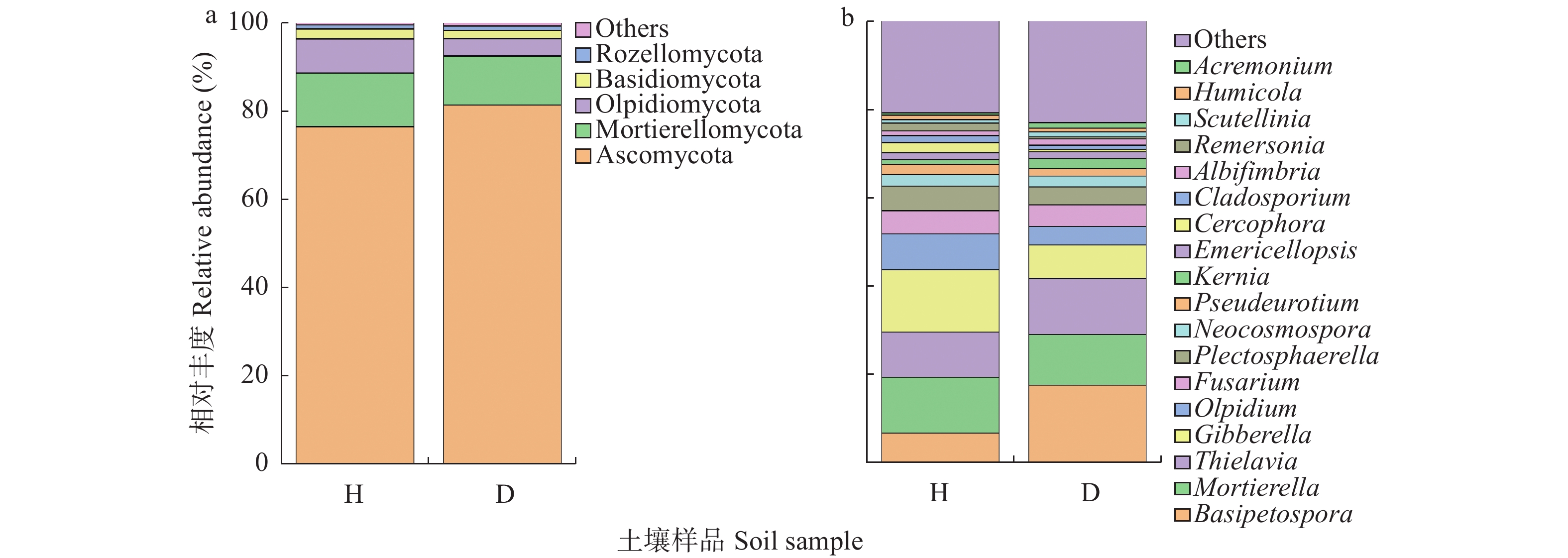
根肿病严重制约着十字花科蔬菜产业的发展, 而土壤微生物多样性与组成的变化不仅与十字花科作物根肿病密切相关, 且对土壤健康、农业生产系统的可持续发展和人体健康至关重要。因此, 了解十字花科根肿病发生与土壤微生物群落结构的关系十分必要。采用Illumina MiSeq高通量测序技术对白菜健康株和患根肿病病株的根围土壤的细菌16S rDNA和真菌ITS序列进行测序, 将结果质控后与相关数据库进行比对; 测定了根围土壤理化性质, 分析了样本间的微生物群落结构与组成差异, 探讨土壤理化、土壤微生物群落和根肿病间的关系; 并对样本细菌和真菌功能进行了预测。结果表明: 1)健康白菜植株根围土壤细菌群落均匀度及多样性程度均高于患根肿病植株, 患根肿病白菜植株根围土壤真菌群落丰富度、均匀性和多样性程度均高于健康白菜植株根围土壤。2)放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)、绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteria)和芽单胞菌门(Gemmatimonadetes)是所测土壤样本中的主要优势细菌门, 芽孢杆菌属(Bacillus)、Gaiella、Defluviicoccus、梭状芽孢杆菌属(Clostridium)和类诺卡氏属(Nocardioides)是所测土壤样本中的主要优势细菌属; 根围土壤优势真菌类群为子囊菌门(Ascomycota)、被孢霉门(Mortierellomycota)、担子菌门(Basidiomycota)和壶菌门(Olpidiomycota), 主要的真菌属为赤霉属(Gibberella)、被孢霉属(Mortierella)、梭孢壳属(Thielavia)和Basipetospora。3)新陈代谢、环境信息处理、细胞过程和有机系统是健康株与患病株根围土壤细菌群落具有显著差异的4类细菌功能; 患根肿病植株根围土壤真菌群落中赤霉属、梭孢壳属、丝壳属(Kernia)和镰刀菌属(Fusarium)的功能丰度均大于健康株。4)主坐标分析结果表明健康植株与患根肿病植株根围土壤细菌和真菌群落结构差异明显, 冗余分析结果显示pH、全氮、碱解氮、速效钾和阳离子交换量是根围土壤微生物群落变化的主要影响因素。白菜健康株与根肿病患病株根围土壤中真菌群落的差异比细菌群落大。本文为研究十字花科作物微生物区系提供了新的依据, 并为研究有益合成菌群抑制病原菌从而控制根肿病的方法提供了可能性, 同时也为缓解土壤退化和重建健康土壤提供了途径。
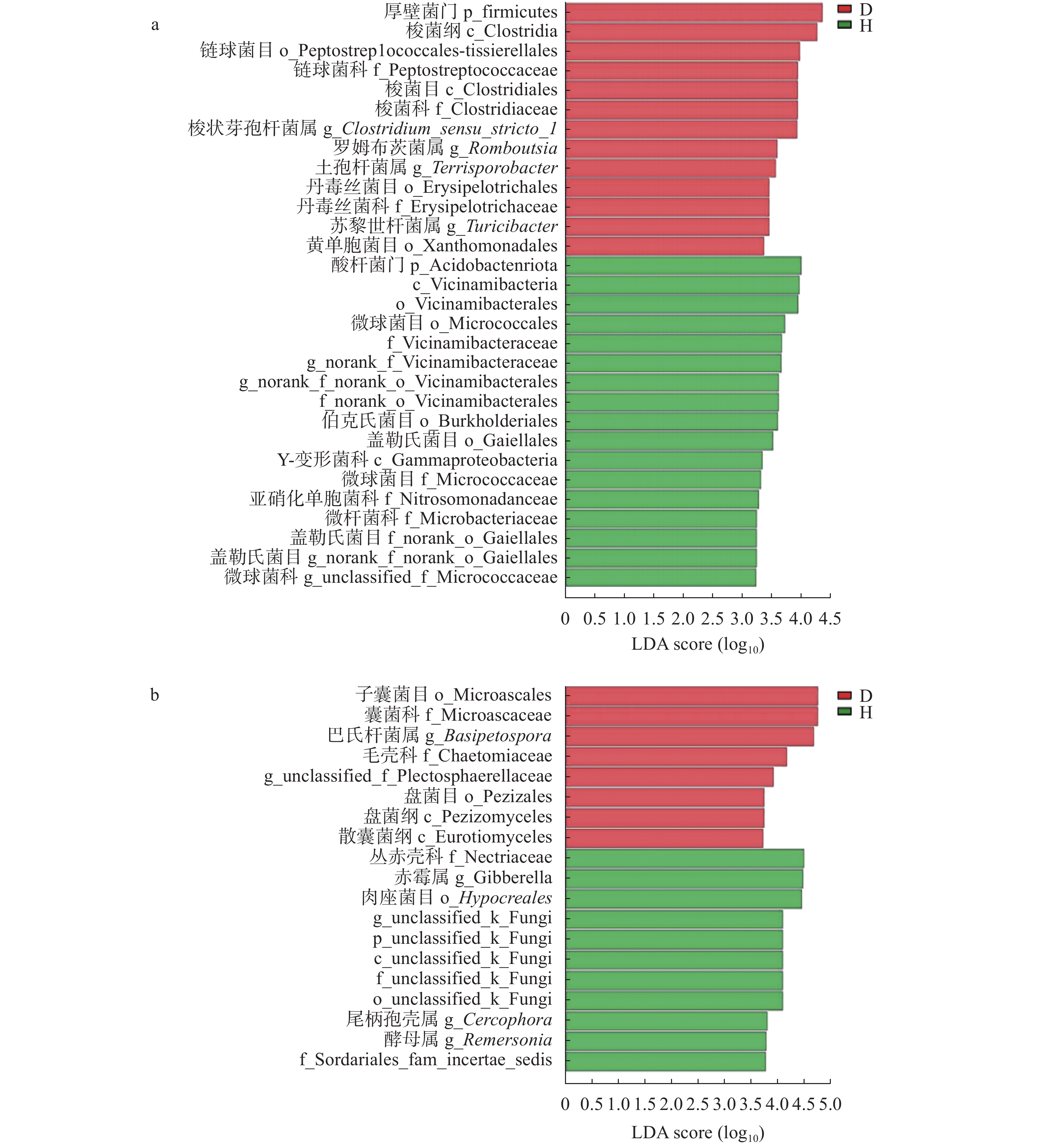
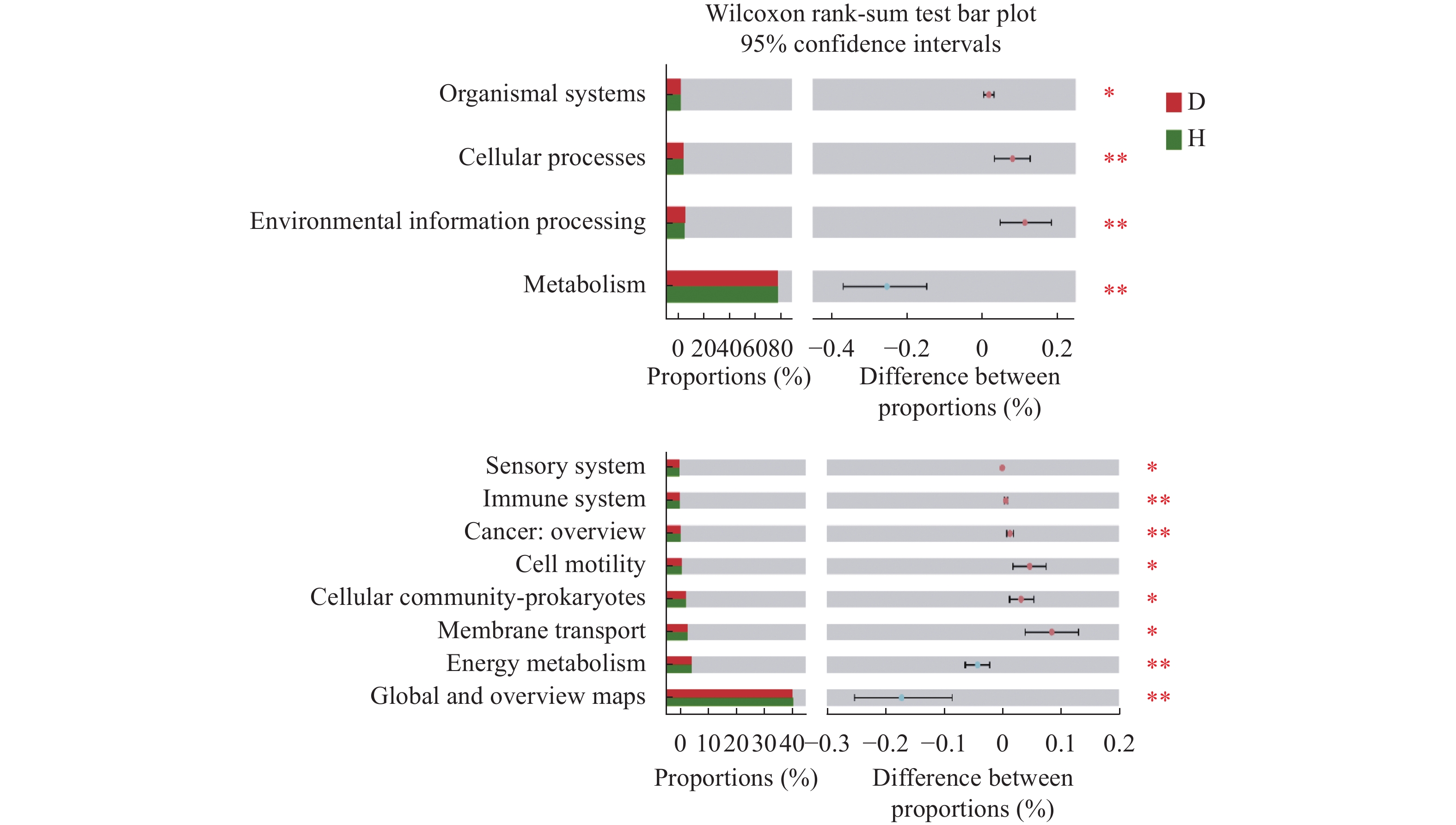

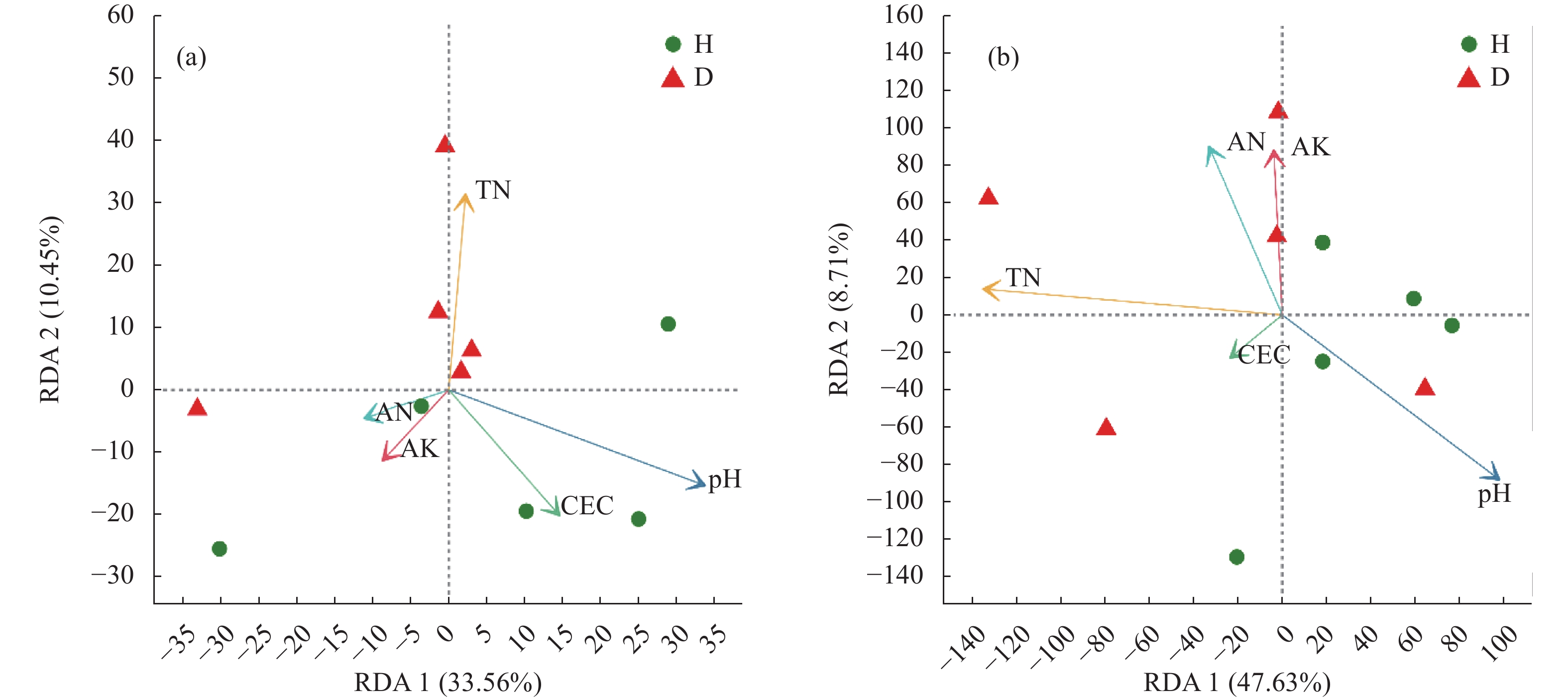
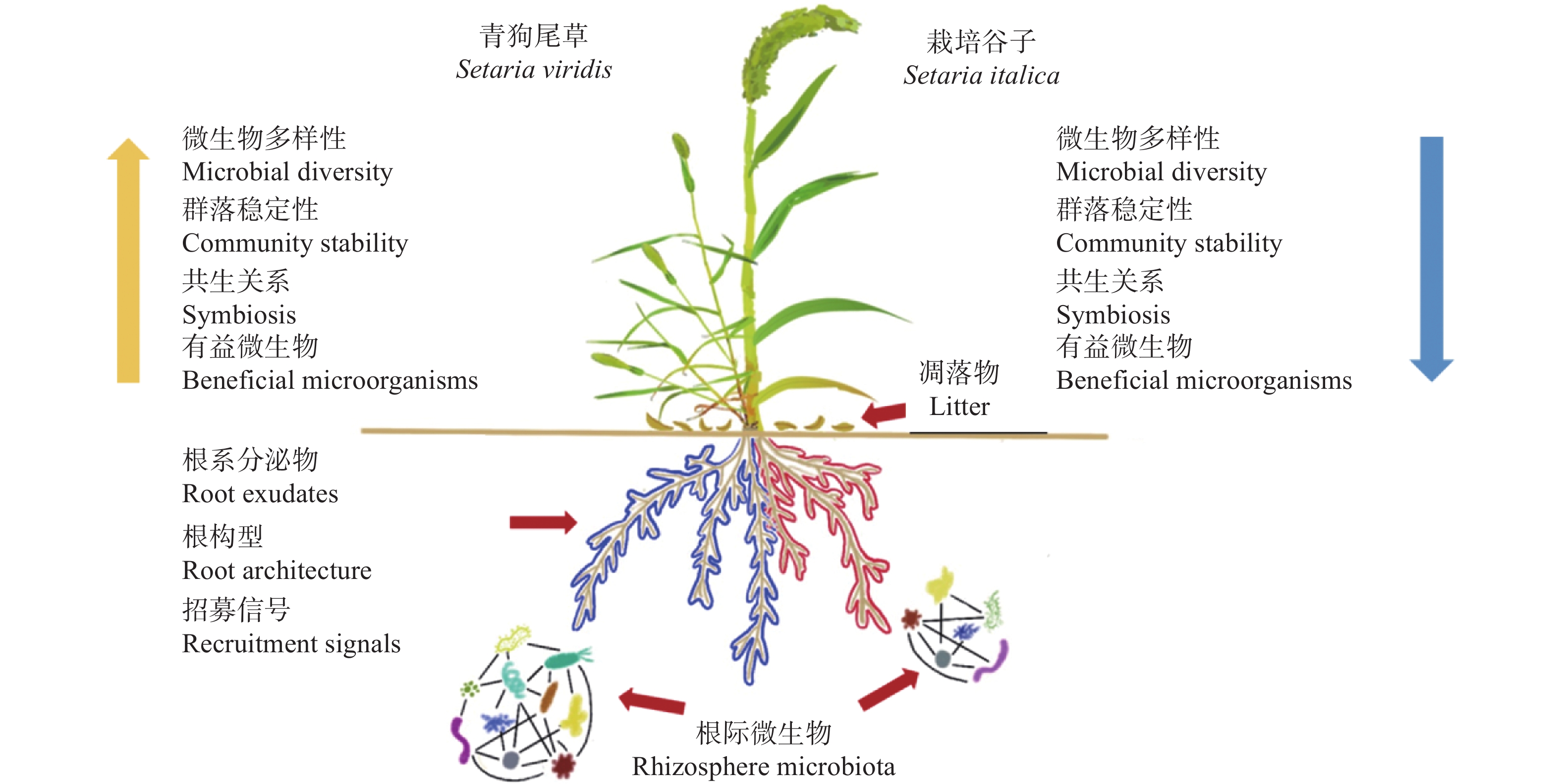
根际微生物可赋予植物新的抗逆性, 其群落结构及生态功能的变化也体现了作物对环境变化的适应能力。驯化影响作物的遗传多样性、生理性状及其代谢产物, 进而对根际微生物群落组成与功能产生作用。目前, 对于作物驯化的研究多关注植物遗传多样性与植物表型和生理特性之间的相互关联及影响机制, 微生物在作物驯化过程中的作用及其与植物互作机制的探索还处于起步阶段。为此, 本文首先对驯化过程在禾本科作物根际微生物群落构建中的作用进行了回顾, 针对起源于中国的重要旱作C4模式作物谷子, 系统总结了谷子的驯化过程及其对谷子生长、生理性状和根际微生物的影响, 以及驯化对谷子根际微生物间互作和谷子与微生物互作的作用机制, 以期为恢复农田生态系统中的作物-微生物有益关系提供科学依据。

生活污泥中富含的重金属限制其资源化利用, 为钝化重金属活性, 降低污泥毒害效果和提高其利用价值, 以40 mL蚯蚓粘液和2%、4%、6%、8%秸秆炭为添加剂对2 kg污泥进行堆肥, 研究粘液、粘液协同秸秆炭添加对污泥堆肥后重金属变化的影响。结果显示, 与对照组污泥堆肥相比, 粘液堆肥污泥后pH升高1.42%, 总氮、总磷含量降低7.87%、14.18% (P<0.05); 而粘液协同秸秆炭堆肥污泥后, 污泥逐渐呈碱性, 电导率提升5.71%~9.58% (P<0.05), 有机质含量升高7.71%~24.60% (P<0.05), 丰富了堆体中可溶性离子和有机物含量, 但总氮、总钾含量分别降低19.10%~30.95%、7.87%~14.31%。在添加粘液对污泥堆肥后, 重金属总量均表现出下降趋势, Ni、Zn、Pb的较活泼形态向难以降解的残渣态转化, 使残渣态所占比例较CK处理分别升高61.81%、120.19%、72.51%; 当添加粘液和秸秆炭对污泥堆肥后, 重金属总量继续表现出下降趋势, 碳酸盐结合态Ni和Pb、铁锰结合态Pb、可交换态Zn逐步向稳定的残渣态转化, 而有机结合态Cu却向可交换态和残渣态转化, 钝化了堆肥污泥中Ni、Zn、Pb, 活化了Cu。最后根据分析得出结论, 粘液协同秸秆炭改变污泥中pH来影响重金属Ni、Zn、Pb、Cu有效态, 粘液+8%秸秆炭处理对污泥重金属的影响较为理想。
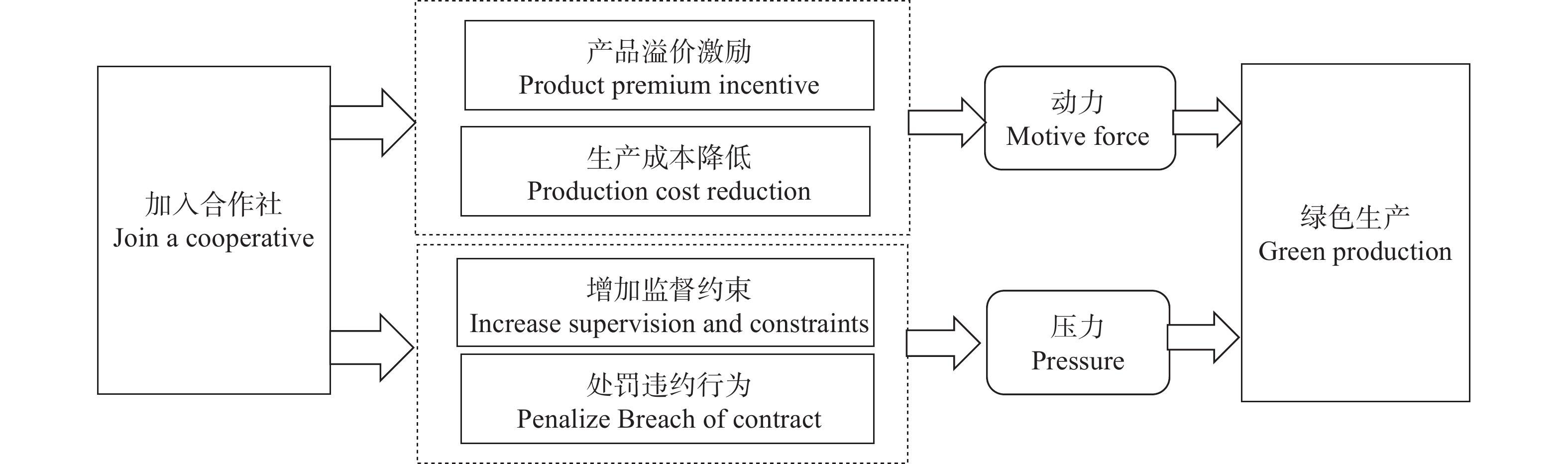
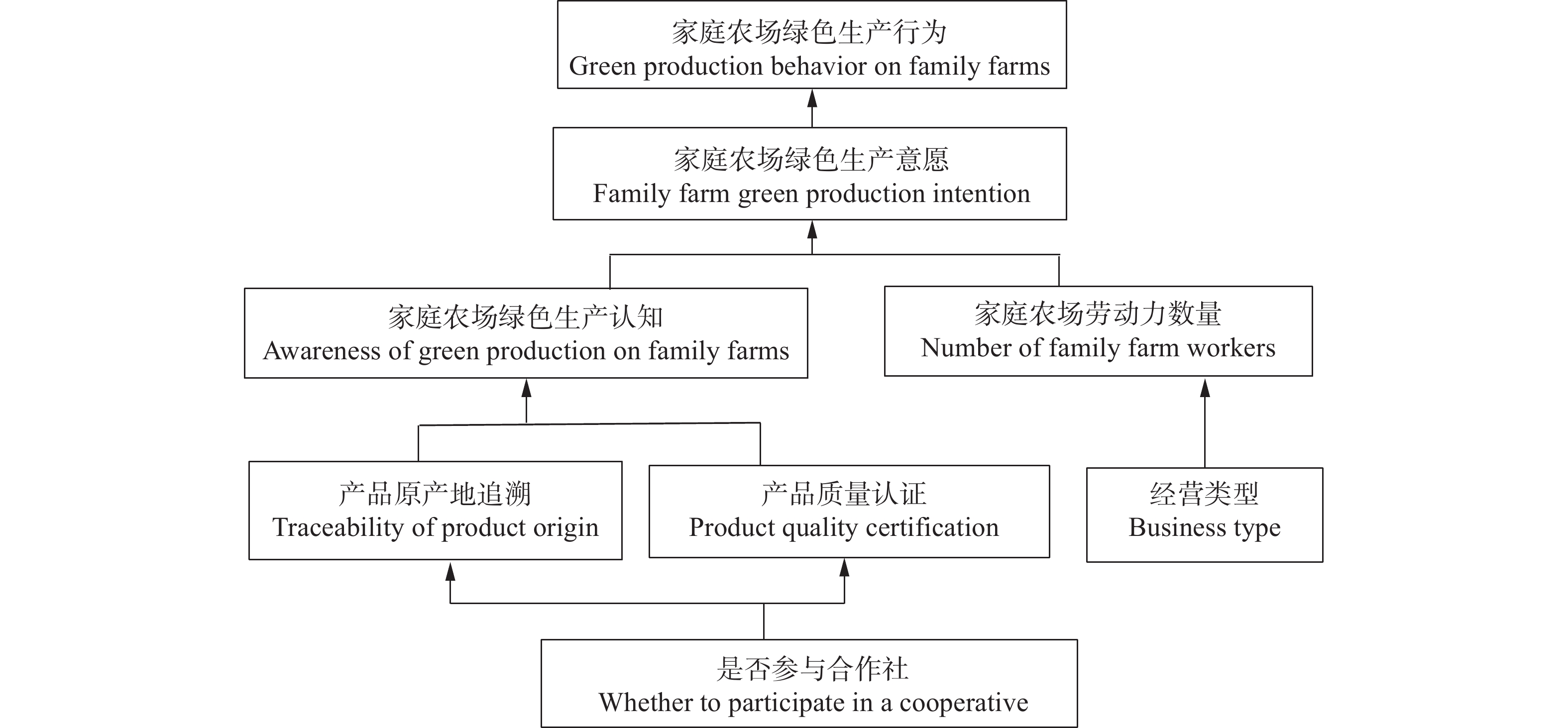
绿色生产是农业可持续发展的必由之路, 家庭农场作为现代农业的关键主体, 探索其绿色生产的有效方式具有重要理论价值和现实意义。本文基于山东省422家省级示范家庭农场的调研数据, 运用排序选择模型和解释结构模型, 实证分析家庭农场加入合作社对其绿色生产的影响。研究发现: 1)加入合作社可以显著促进家庭农场绿色生产, 且对节水灌溉、施用有机肥、水肥一体化的促进作用更明显。2)不同农场加入合作社后的绿色生产行为存在明显差异, 作为领办人的农场要优于作为普通社员的农场, 粮食、蔬菜类农场显著, 中等规模(10~20人)农场显著。3)家庭农场的经营类型、劳动力数量、产品质量认证、原产地追溯、绿色生产认知和意愿均对其绿色生产产生影响。4)解释结构模型发现, 加入合作社作为深层根源因素直接影响家庭农场绿色生产认知, 并进而影响绿色生产意愿, 最终促成绿色生产行为。为此, 应鼓励并支持家庭农场加入合作社, 重视农场间的异质性, 通过合作社的培训与监督促进农场绿色生产。

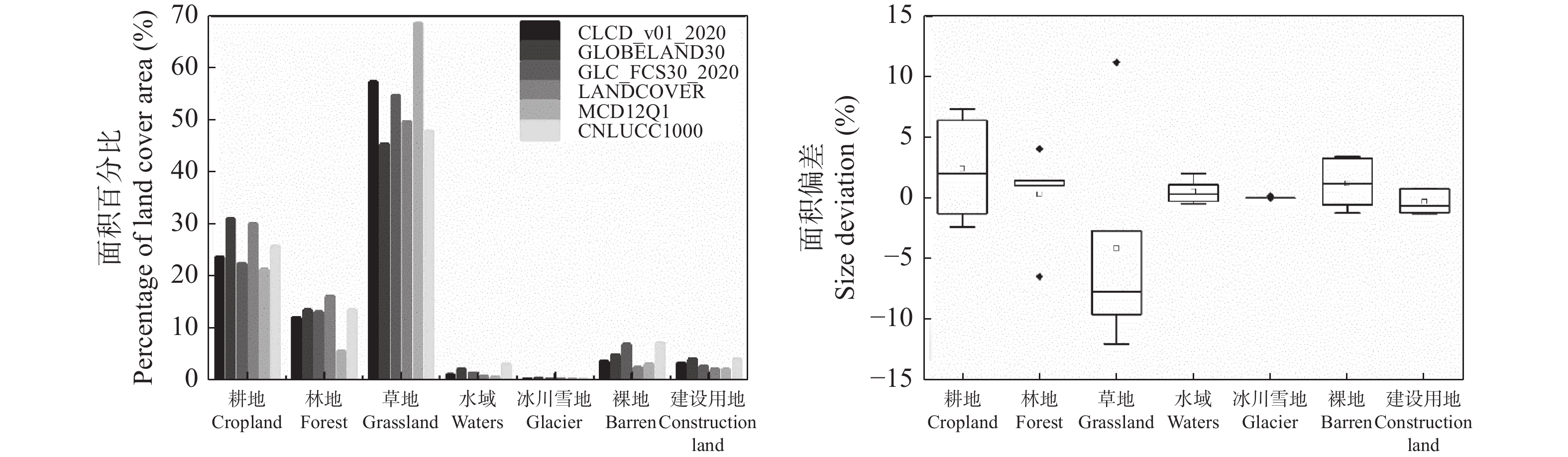
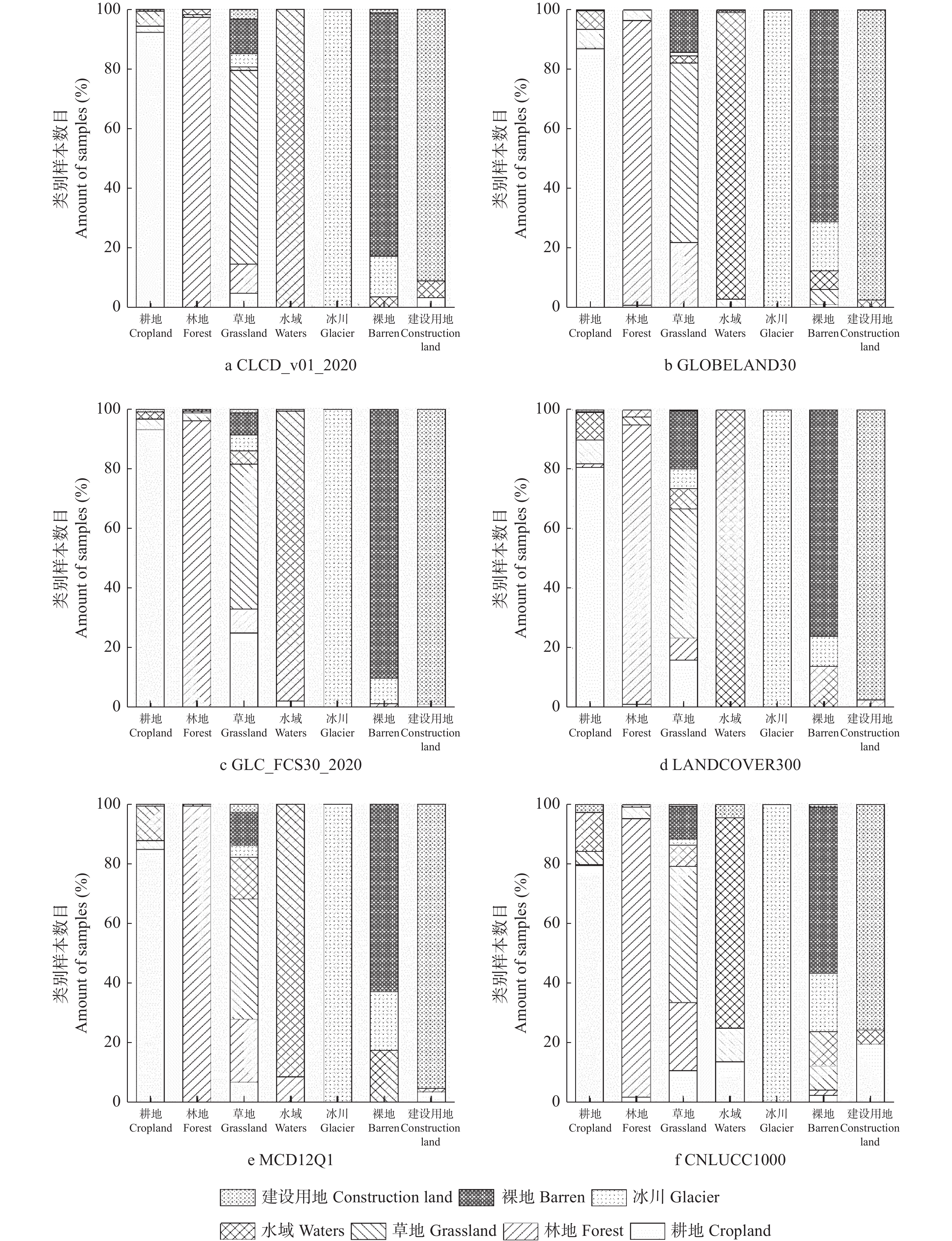
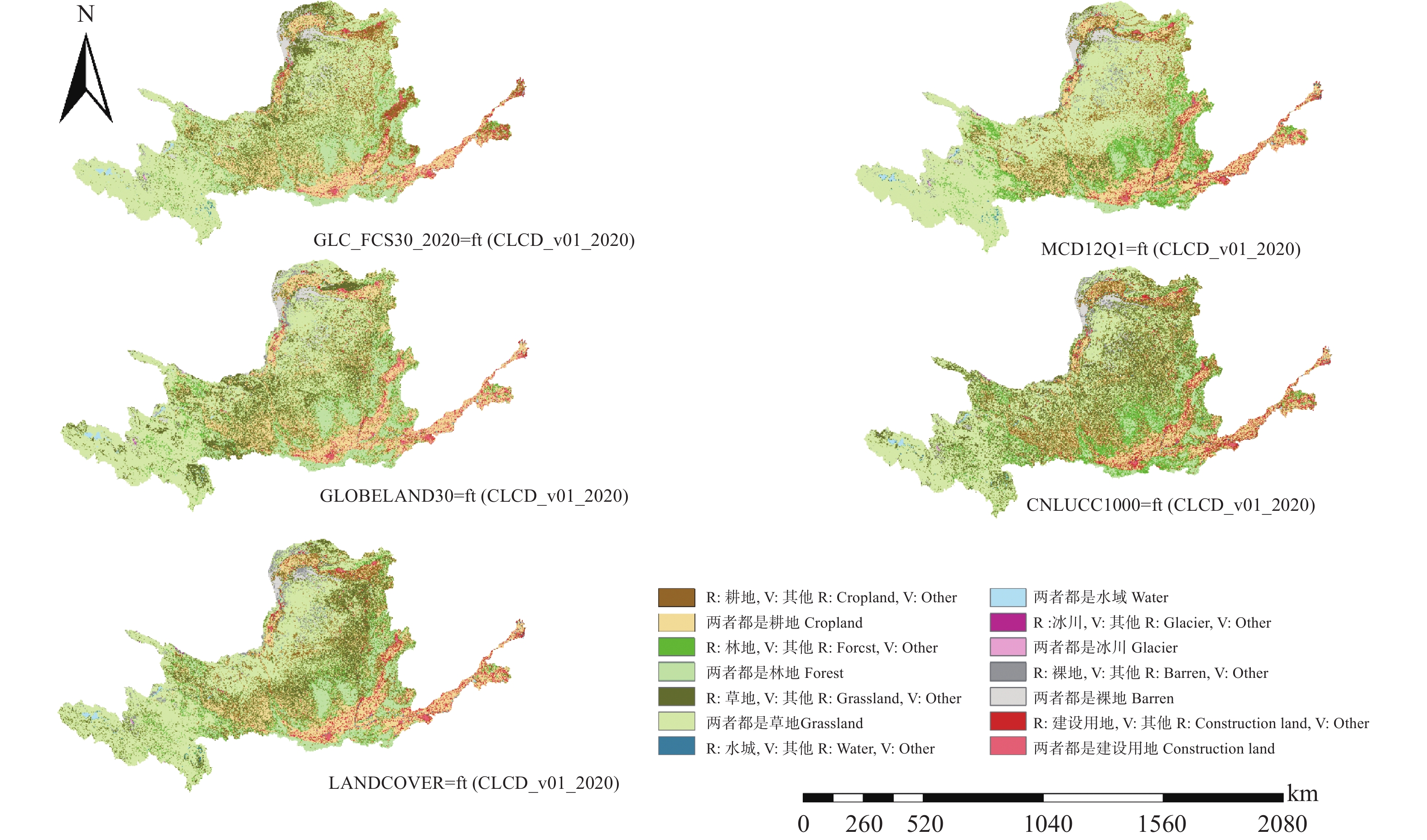
开源、多分辨率、及时的土地覆盖产品为了解全球地表覆盖状况、陆面过程模型模拟以及社会经济发展决策等提供了丰富的数据支撑, 但多源的数据存在不同程度的不确定性, 在区域尺度如何选择合适的土地覆被产品成为应用中的难题。本研究以黄河流域为例, 对分辨率从30 m到1000 m的CLCD_v01_2020、GLOBELAND30、GLC_FCS30_2020、LANDCOVER(300 m)、MCD12Q1(500 m)和CNLUCC1000(1000 m)等6种2020年土地覆被产品进行区域尺度精度评价和一致性分析。基于Google Earth采集的1540个样本点分析6种数据在黄河流域的总体精度, 以最高精度的数据为参考对其他数据进行面积一致性分析, 并对6种数据进行类别混淆分析和混淆图谱分析。结果表明, 6种数据中分类精度最高的为CLCD_v01_2020, 总体精度(overall accuracy, OA)达88.12%; 其次是GLOBELAND30 (OA=85.32%)、GLC_FCS30_2020 (OA=84.09%)、LANDCOVER300 (OA=77.79%)、MCD12Q1 (OA=73.38%)、CNLUCC1000 (OA=71.82%), 30 m土地覆被产品的KAPPA系数均在0.8以上, 随着空间分辨率的下降, 分类精度下降。 6种数据的土地覆被类别组成的相对比例总体上趋于一致, 但在耕地和草地两类土地覆被类别上仍存在较大差异, GLC_FCS30_2020与参考数据CLCD_v01_2020的相关性最高, R2达到0.9976。通过类别混淆分析可知6种数据普遍对耕地、林地和草地的混淆较为严重。类别混淆空间分析表明, 验证数据与参考数据在黄河上游的草地, 中下游部分耕地和建设用地等类型较为单一的区域一致性较高, 而在陕西北部、山西西部的一致性较差, 主要表现为草地和林地的混淆。针对黄河流域土地覆被数据一级分类, 本研究建议, 30 m分辨率的数据中选择CLCD_v01_2020, 百米级分辨率数据中选择LANDCOVER300, 二级分类则可以根据所需的分类体系选择合适的数据。
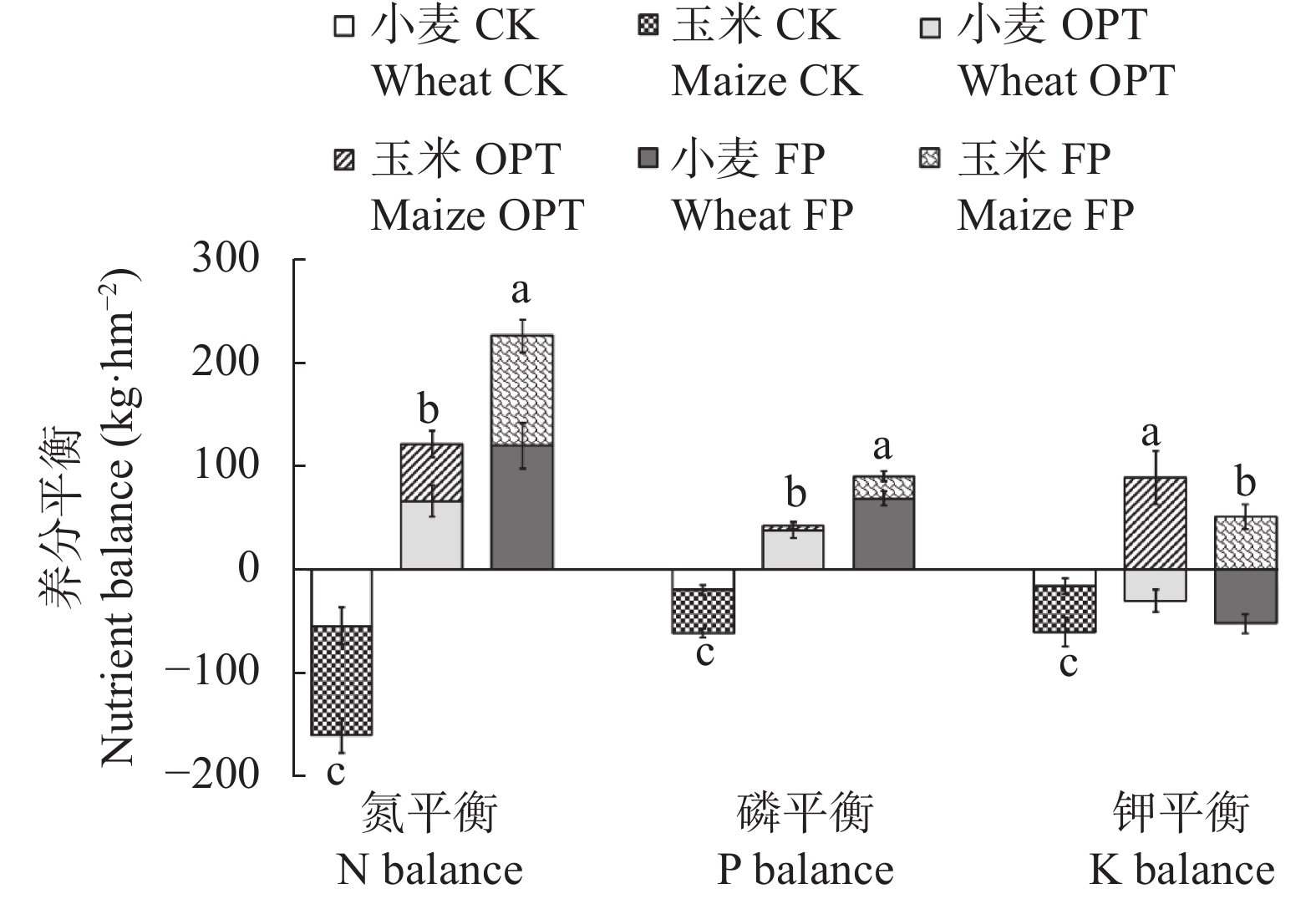
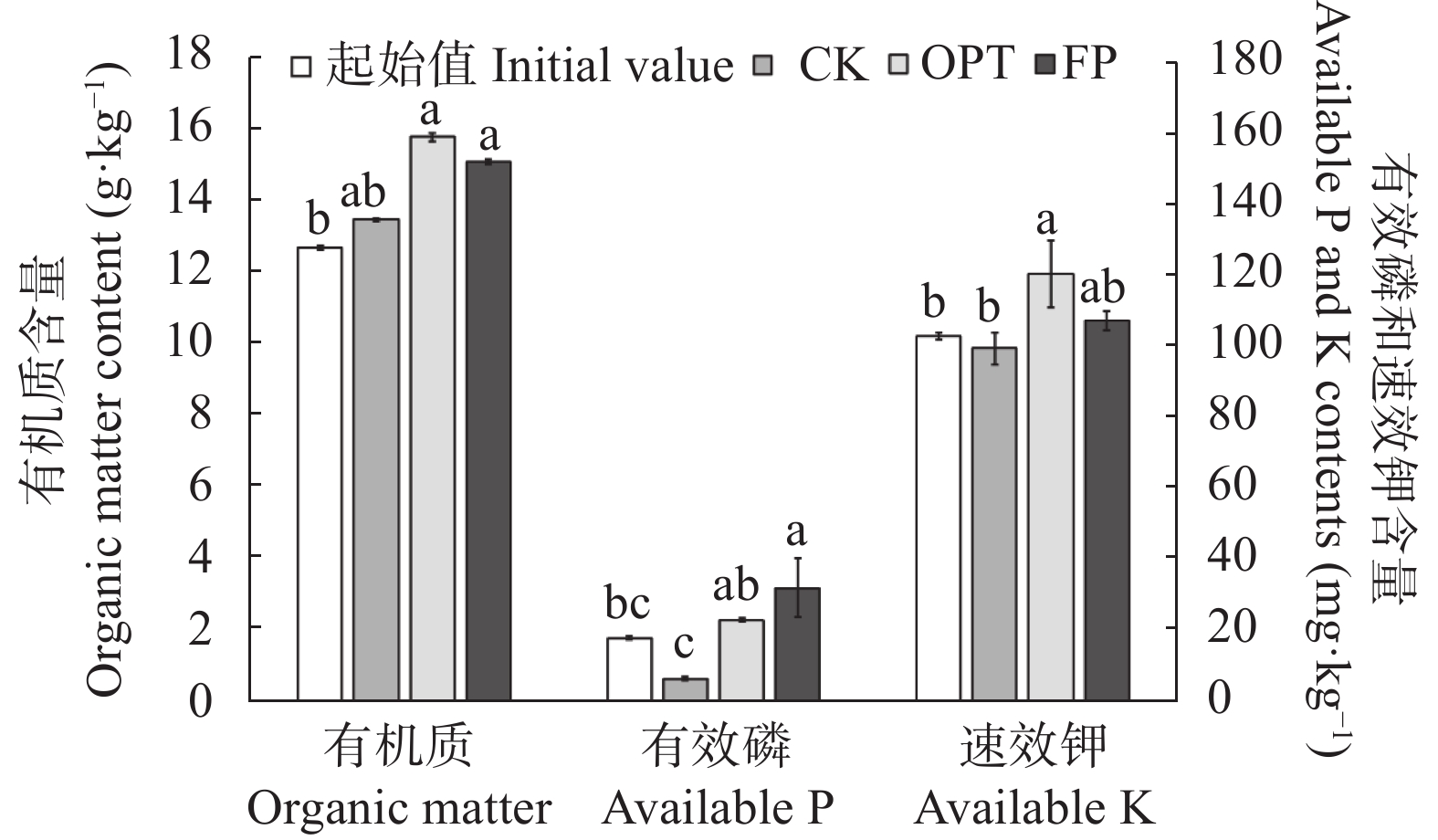
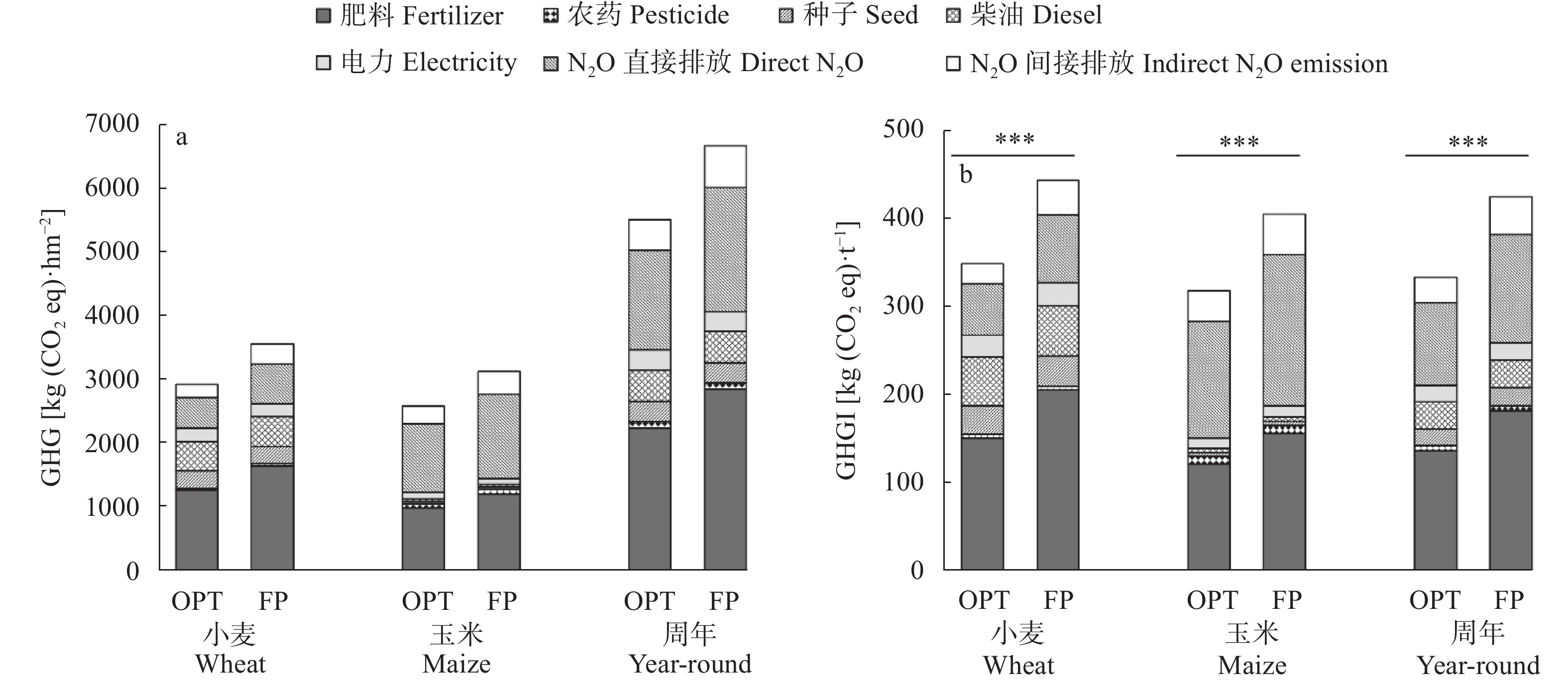
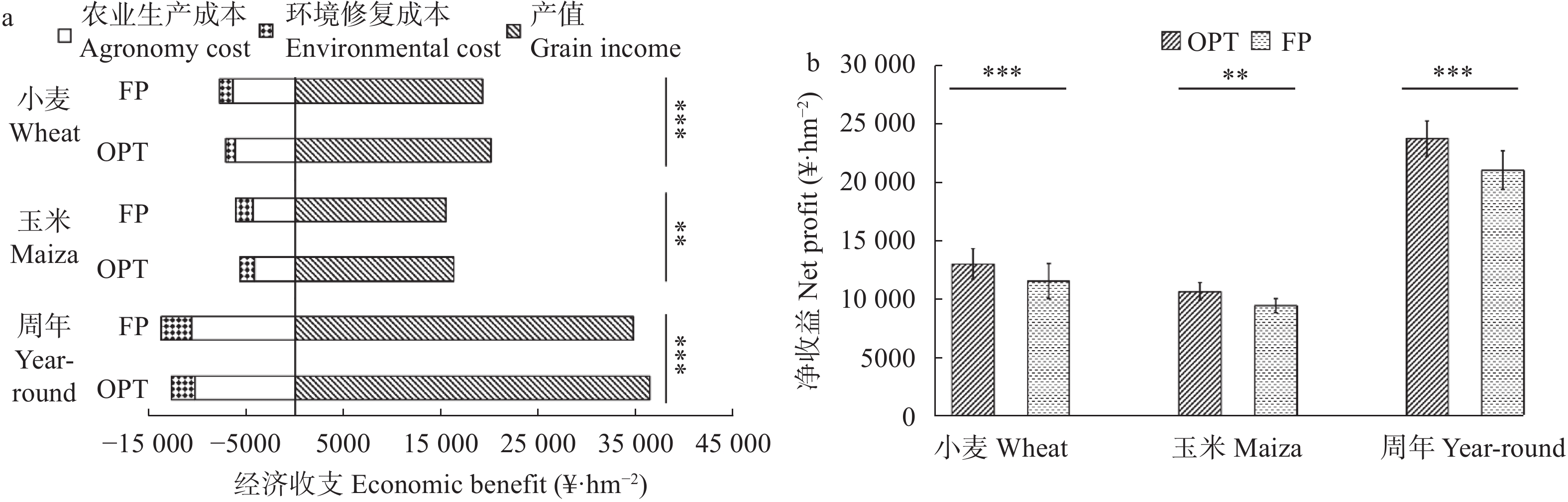
小麦-玉米轮作是华北平原主要的种植模式, 对保障我国粮食安全起着关键作用。本文系统研究了不施肥(CK)、优化施肥(OPT)和农户习惯施肥(FP)方式连续8年对小麦-玉米轮作体系产量、养分平衡、温室气体排放和经济效益的影响。结果表明, 小麦季、玉米季和周年轮作中, OPT较FP产量分别提高4.3%、5.3%和4.8%; 氮肥偏生产力分别提高39.1%、31.7%和35.9%; 磷肥偏生产力分别提高39.1%、40.4%和39.8%; 钾肥偏生产力分别降低47.8%、47.3%和47.6%; 温室气体排放量分别降低21.7%、21.1%和21.4%; 温室气体排放强度分别降低27.0%、27.5%和27.3%; 净收益分别提高11.2%、11.4%和11.3%, 农业生产成本分别降低3.7%、2.1%和3.1%, 环境修复成本分别降低28.4%、17.3%和22.1%。周年轮作中, OPT较FP氮素盈余量降低105 kg·hm−2 (46.3%); 磷素盈余量降低48 kg·hm−2 (53.3%); 钾素从亏缺1 kg·hm−2变为盈余59 kg·hm−2, 满足了作物生长需求。连续8年, OPT较FP土壤有机质含量提高5.3%, 速效钾提高12.3%, 有效磷降低27.8%。综上所述, 与农户习惯施肥(FP)相比, 优化施肥(OPT)具有高产、高收益以及环境友好的优势, 为华北平原小麦-玉米轮作体系高效绿色生产提供了科学依据。
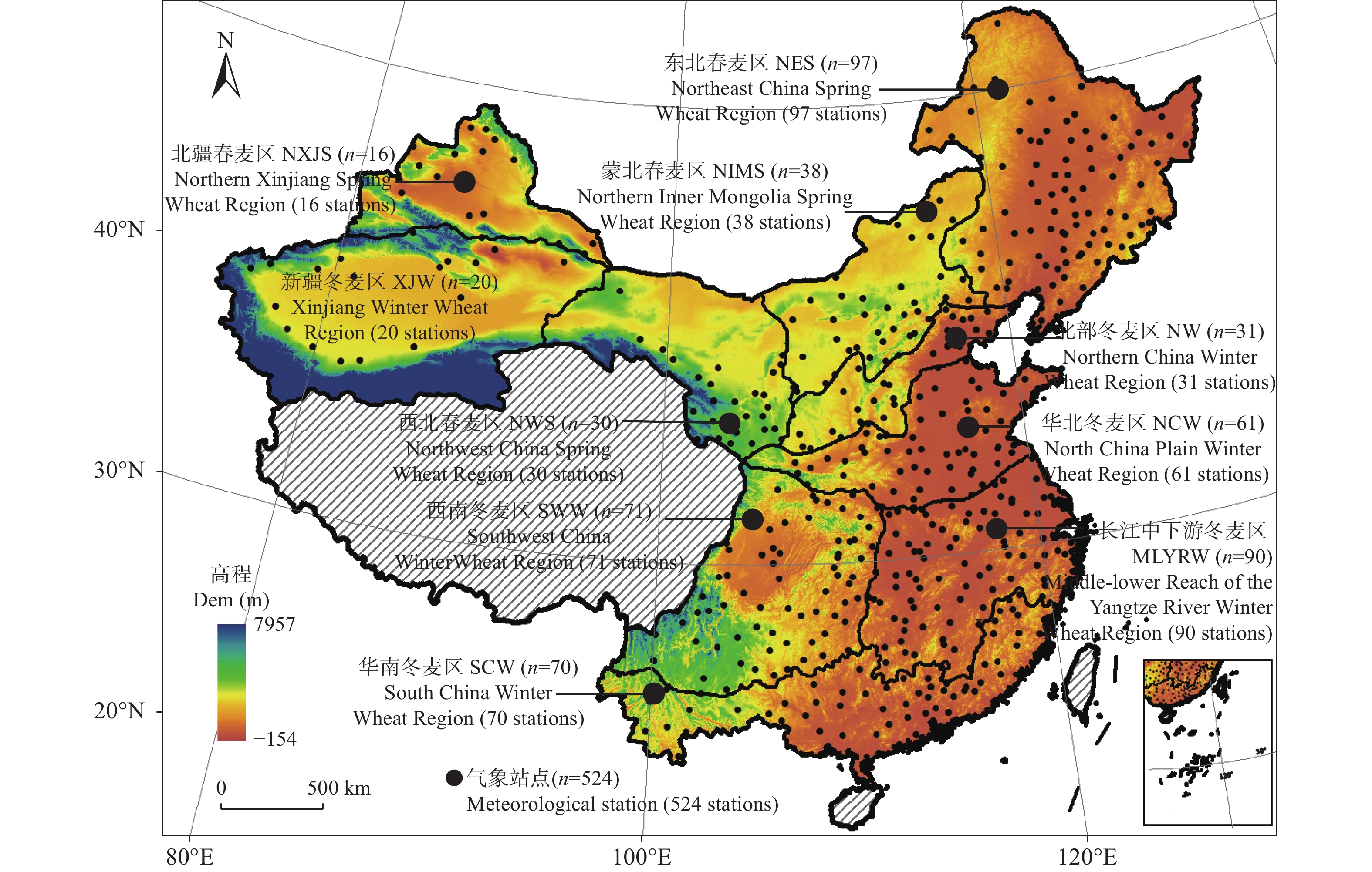
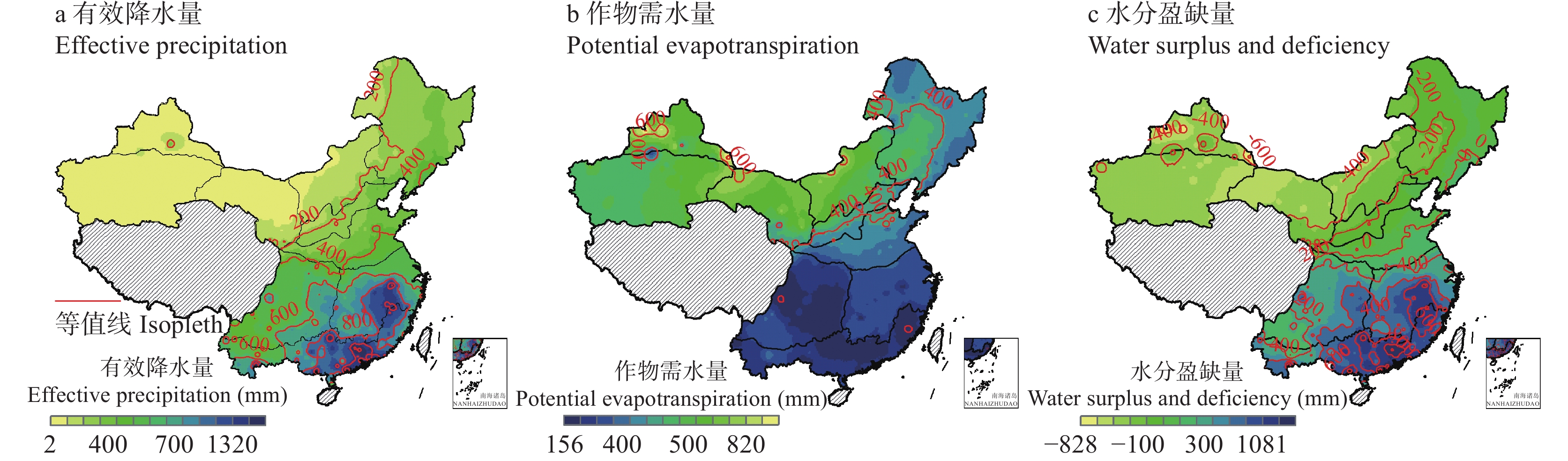
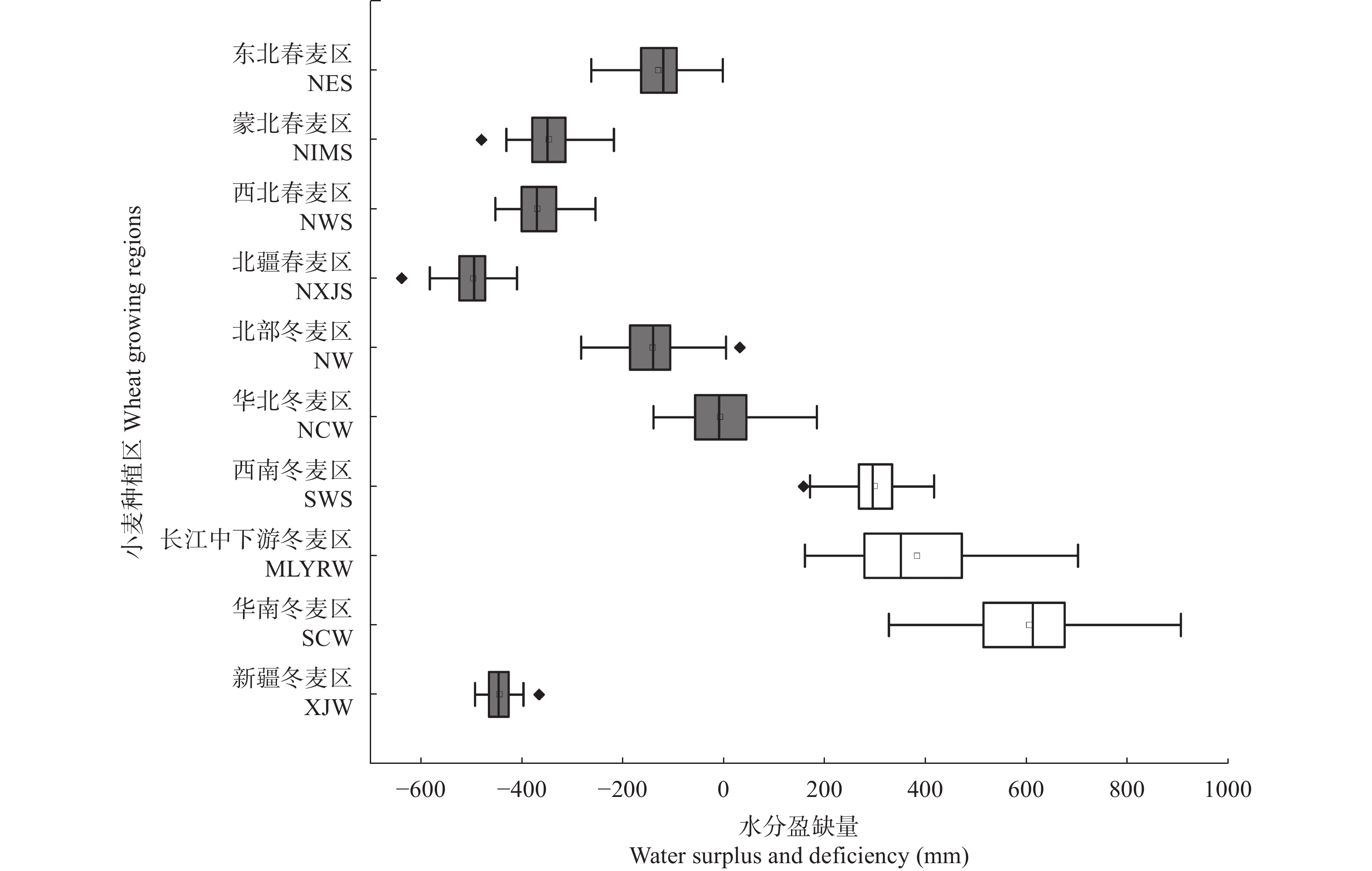
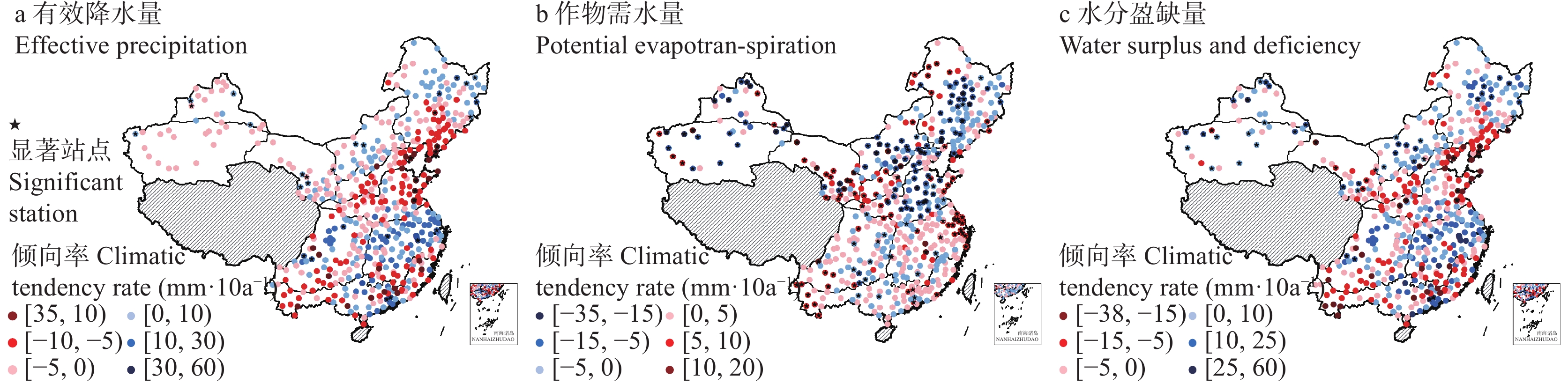
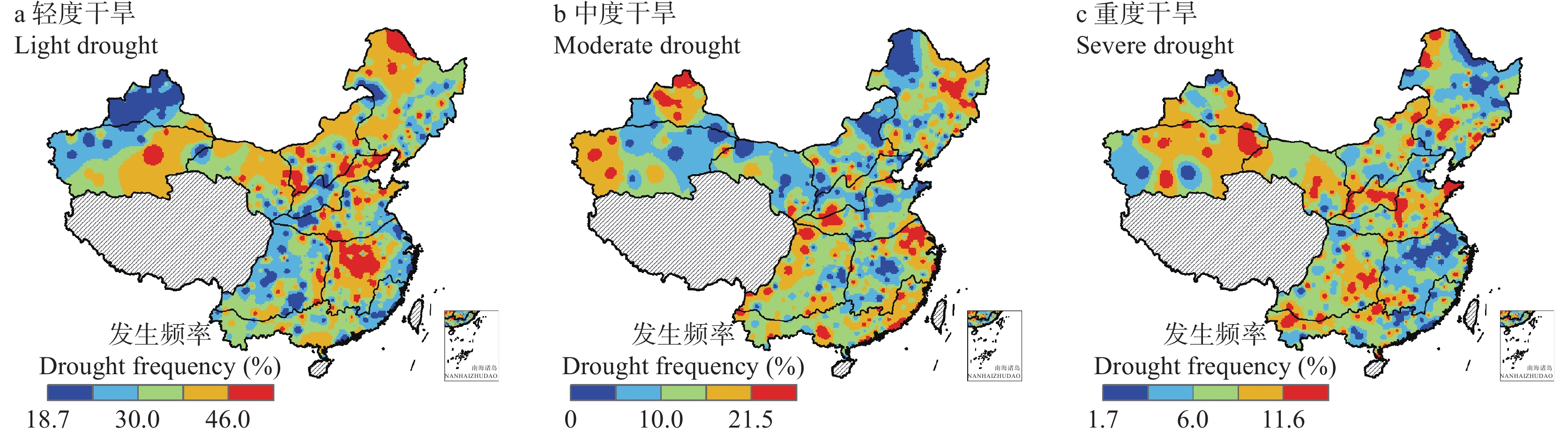
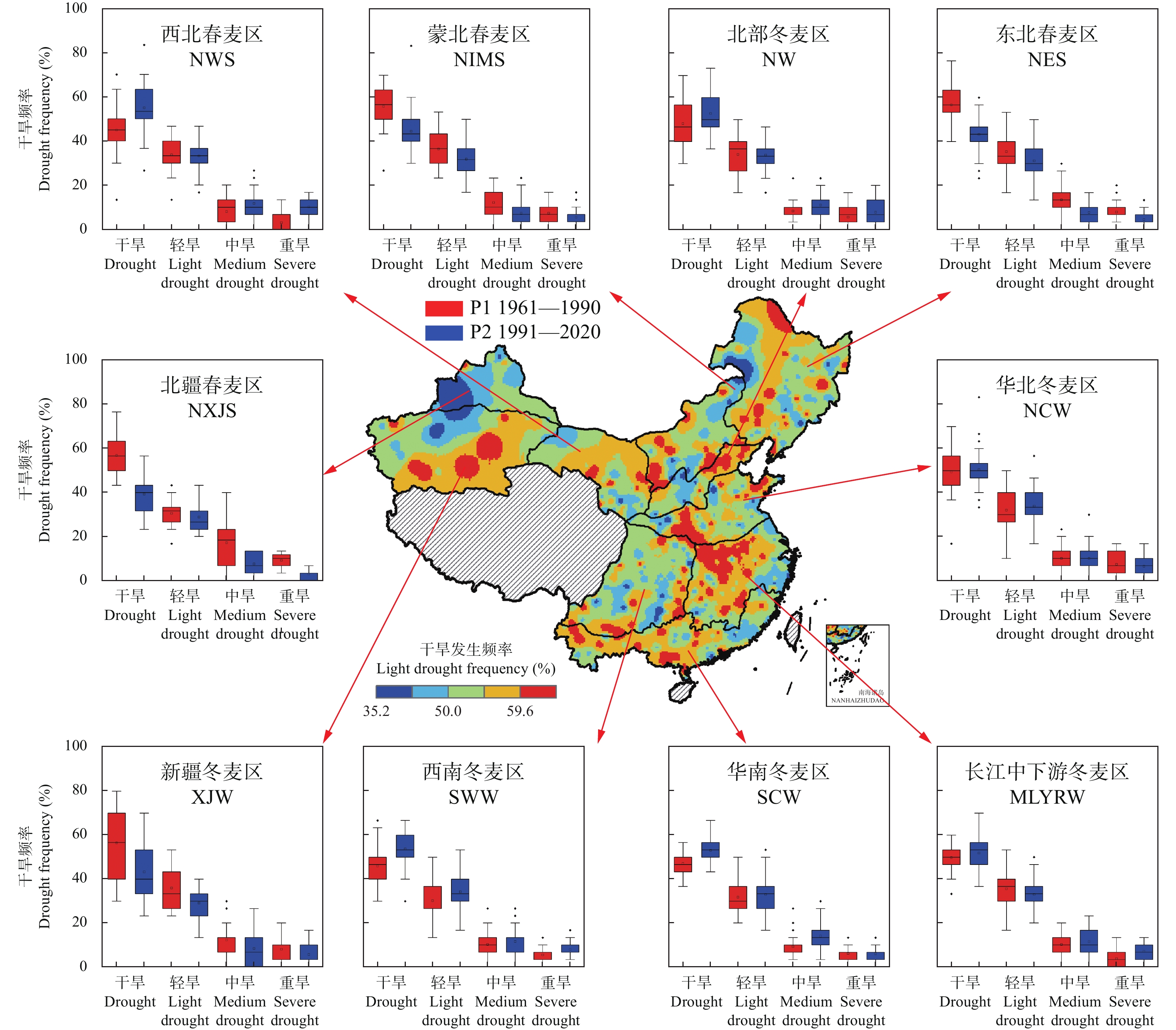
基于1961—2020年全国524个气象台站逐日数据, 以有效降水量、作物需水量和水分盈缺量(有效降水量与作物需水量的差值)为干湿指标, 从年际和年代际尺度(P1: 1961—1990年, P2: 1991—2020年)分析了全国小麦主产区(春麦区: 东北、蒙北、西北、北疆; 冬麦区: 北部、华北、西南、长江中下游、华南、新疆)生长季内气候干湿状况时空分布和演变趋势, 并利用SPEI指数评估了小麦种植区的干旱风险。结果表明: 1) 1961—2020年华南、长江中下游、西南冬麦区小麦生长季降水量大于作物需水量, 其他麦区生长季内水分亏缺, 新疆冬麦区(443 mm)和北疆春麦区(495 mm)为缺水量高值区。2)近60年全国小麦生长季干旱频率为35.2%~59.6%, 四大春麦区、长江中下游冬麦区干旱发生频率较高, 均大于50.0%。3) 1961—2020年全国小麦生长季有效降水量波动增加, 作物需水量呈先降后升的趋势, 华北、北部冬麦区表现为气候暖干化, 其他麦区均呈气候暖湿化趋势。小麦种植区暖湿化的气候机制存在地域间差异, 春麦区(东北、蒙北、北疆)和新疆冬麦区为有效降水量增加且作物需水量减少; 而长江中下游冬麦区生长季内有效降水量和作物需水量均为增加的趋势, 但降水量增加幅度大于作物需水量。本文在全国尺度上对全国小麦生长季干湿状况时空变化进行了探究, 对农业正确应对气候变化具有重要的参考意义。
科学评价区域内大豆生产的生态效率, 有利于促进区域内大豆产业的可持续发展。本研究以华北平原大豆生产典型县——石家庄市藁城区50个农户为例, 基于生命周期评价法(LCA)和超效率(SBM)模型, 对其进行了生命周期评价和生态效率分析。结果显示, 大豆生产4个主导的潜在环境影响类别依次为全球变暖潜力(global warming potential, GWP)、陆地生态毒性潜力(terrestrial eco-toxicity potential, TETP)、酸化潜力(acidification potential, AP)及富营养化潜力(eutrophication potential, EP)。其中, 种植规模方面, 大规模农户的GWP、TETP及EP影响潜力最大; 生态效率值为大规模>中规模>小规模; 其6个投入指标当中, 杀虫剂的冗余率极差最大(5.89%)。灌溉模式方面, 滴灌的GWP和AP影响潜力最大, 沟灌的TETP和EP影响潜力最大; 生态效率为滴灌>喷灌>无灌溉>沟灌; 6个投入指标中, 灌溉用水的冗余率极差最大(8.40%)。种植区域方面, 藁城北部地区的GWP、AP和EP影响潜力均大于藁城南部地区; 生态效率值为藁城南部地区>藁城北部地区; 6个投入指标中,化肥的冗余率极差最大(2.79%)。综上所述, 藁城区大豆生产应向大规模化发展, 并积极推广滴灌技术, 控制化肥和杀虫剂使用量, 以保证大豆产量的同时, 提高大豆生产的生态效率。研究结果可为藁城区大豆生产的生态评价提供参考依据, 有助于其大豆产业的可持续发展。
青海省草地资源丰富, 但受人口增长、气候变化等因素的影响,草地面临大规模退化。由于传统、文化、生态等因素,高寒牧区牲畜出栏时间普遍较晚,这种传统的畜牧业生产经营模式导致了草地超载, 进一步加剧了草地退化。为探寻出兼顾经济效益与生态效益的畜牧方案, 本研究以青海省果洛藏族自治州玛多县为例, 基于实地调研收集的2012—2021年畜牧业统计数据与牧户信息, 根据IPCC提供的家畜能量需求计算方法估算发现,玛多县近10年畜牧业生产消耗的牧草量为44.30万吨/年,与其草地生态承载量37.76万吨/年相比超载了17.34%。其中,由于牲畜出栏晚导致的牧草浪费达到13.69万吨/年,占牧草消耗量的30.90%。本研究进一步设置了“牲畜出栏量不变-提前出栏”和“牲畜存栏量不变-提前出栏”两种假设优化情景, 计算了不同优化情景下的牧草消耗量和肉产量。结果显示,若提早牲畜出栏年龄, 可将玛多县平均草料转化率(即每万吨牧草干物质可生产的肉产量)较当前水平提高23%~86%;同时,在不超过草地生态承载力的前提下, 肉产品增产潜力相比当前水平最高可达58%,可提高畜牧业产值6226万元。
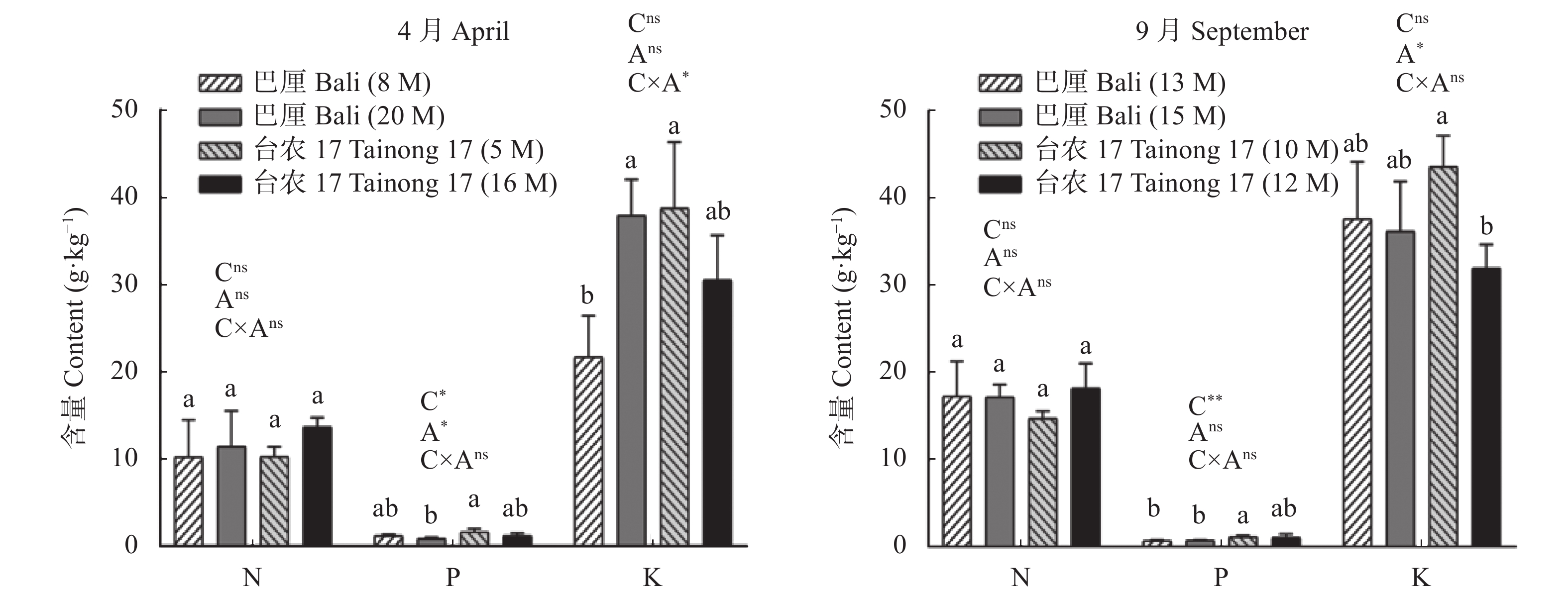
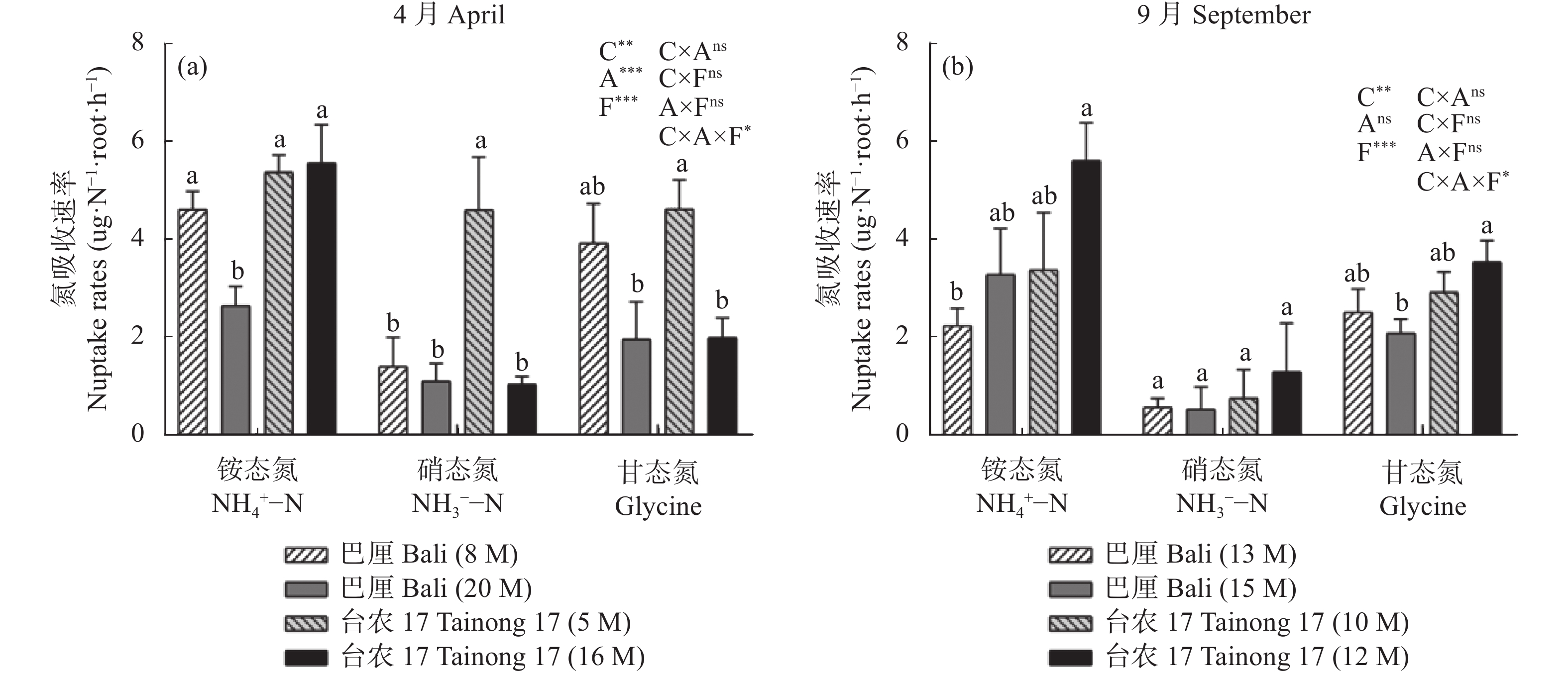
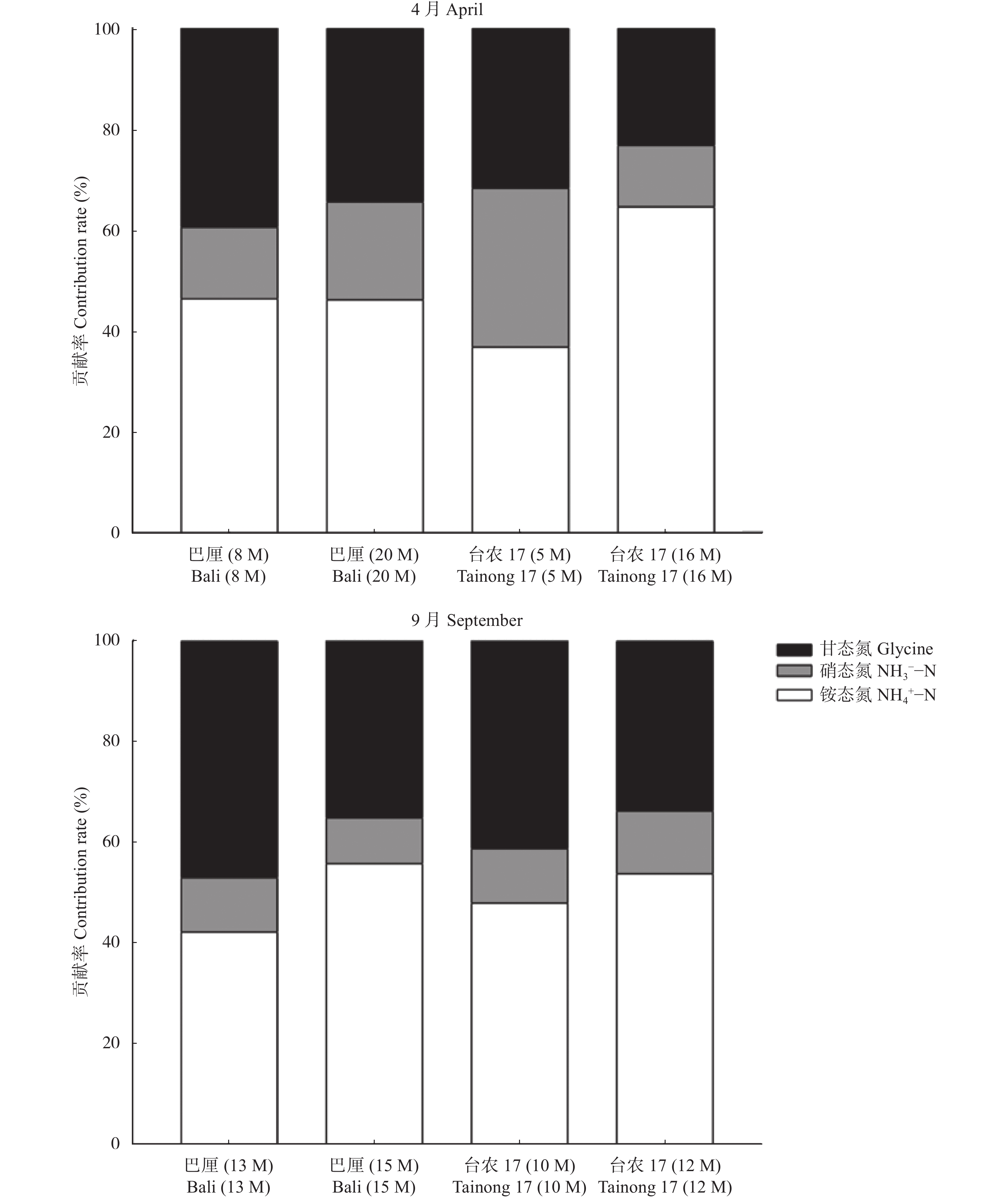
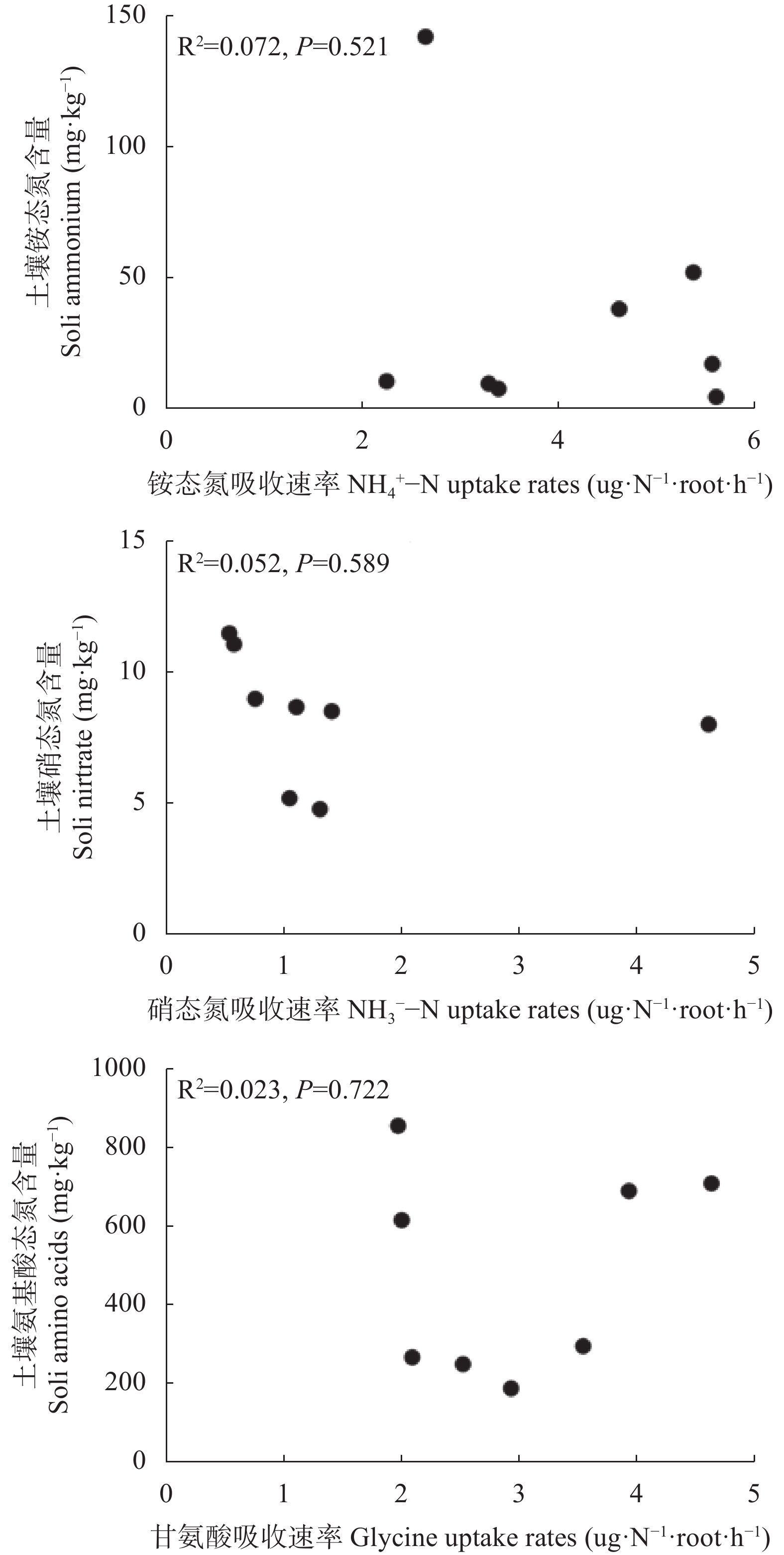
氮是菠萝最需要的大量营养素之一, 也是与产量关系密切的营养元素。本试验分别在4月和9月两个生长季节, 选择了广东省徐闻县田间生长的‘巴厘’和‘台农17’两个菠萝品种不同树龄的植株为研究对象, 测定了不同年龄植株的形态、生理和生长特征, 并利用稳定性同位素15N示踪技术探讨了菠萝对3种形态氮素(铵态氮、硝态氮和甘氨酸)的获取策略。结果表明, 在4月份果实收获期, 与‘巴厘’相比, ‘台农17’菠萝的产量(单个鲜果重)和根生物量较低, 但其植株高度、单株生物量、叶片N和K含量和比叶面积无显著差异, 叶片碳稳定性同位素(δ13C)和P含量较高。无论在4月份或者9月份, 两菠萝品种间对不同形态的氮素吸收有显著的差异。总体而言, ‘台农17’比‘巴厘’的氮吸收能力要强(P<0.05)。‘台农17’菠萝较强的氮吸收能力和水分利用效率更有助于将其分配到地上以促进光合作用, 从而维持其植株在较短生命周期内的生长。两菠萝品种都偏好吸收铵态氮(36.8%~64.6%), 其次是甘氨酸(23.2%~47.1%), 对硝态氮吸收速率最低(9.1%~31.5%)。处于营养生长阶段的菠萝植株(5~8个月)比果实收获时期的氮吸收速率高。随着树龄的增长, 铵态氮贡献率逐渐增大, 而甘氨酸贡献率逐渐降低。不同季节和树龄条件下, 不同形态氮素的吸收速率与土壤氮含量和其他所测得植物性状的相关性不显著。总之, 本研究首次证实田间菠萝的根系具有较强直接吸收利用有机氮的能力, 菠萝的品种和生长阶段都是影响氮素获取策略的重要因素。

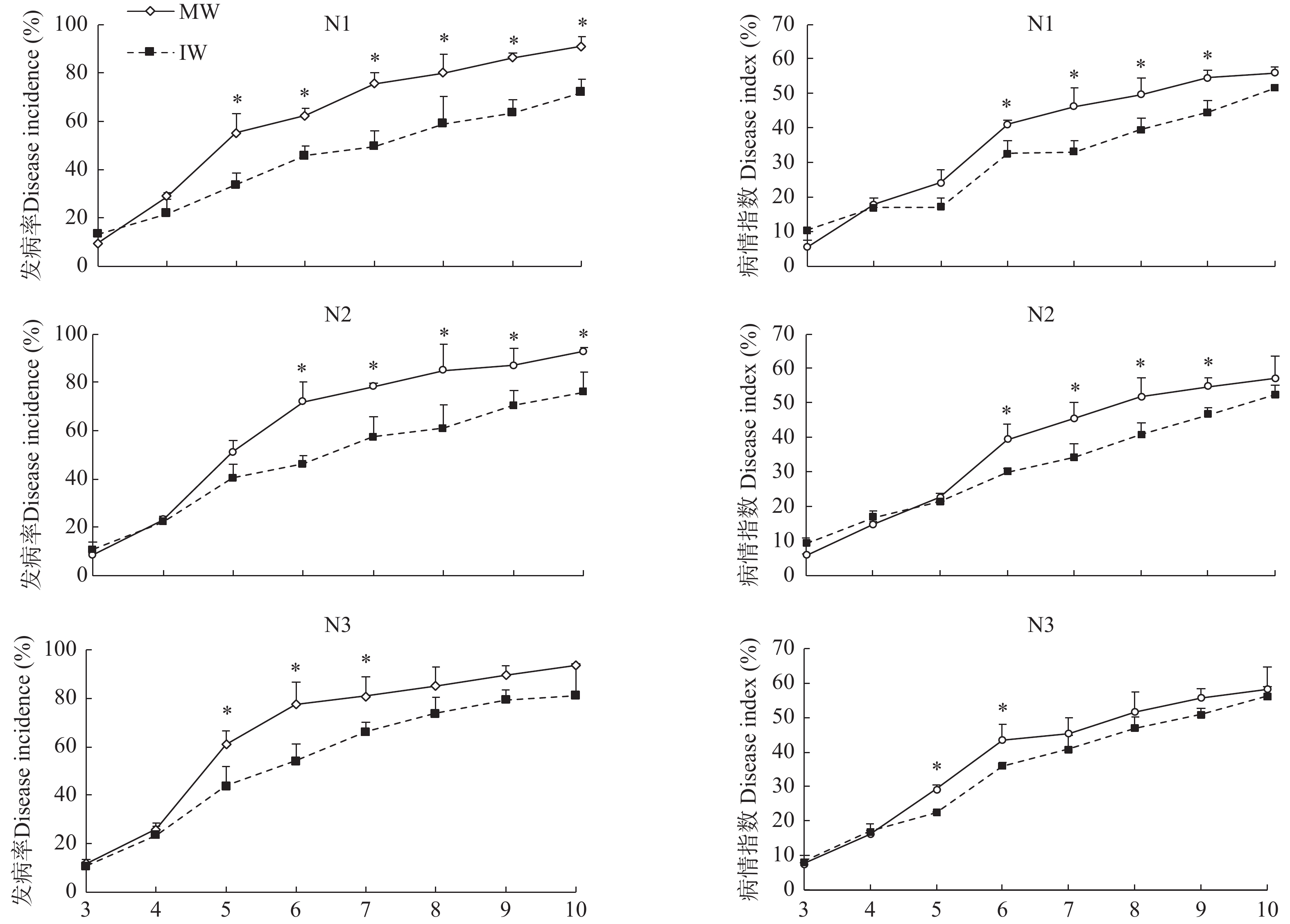
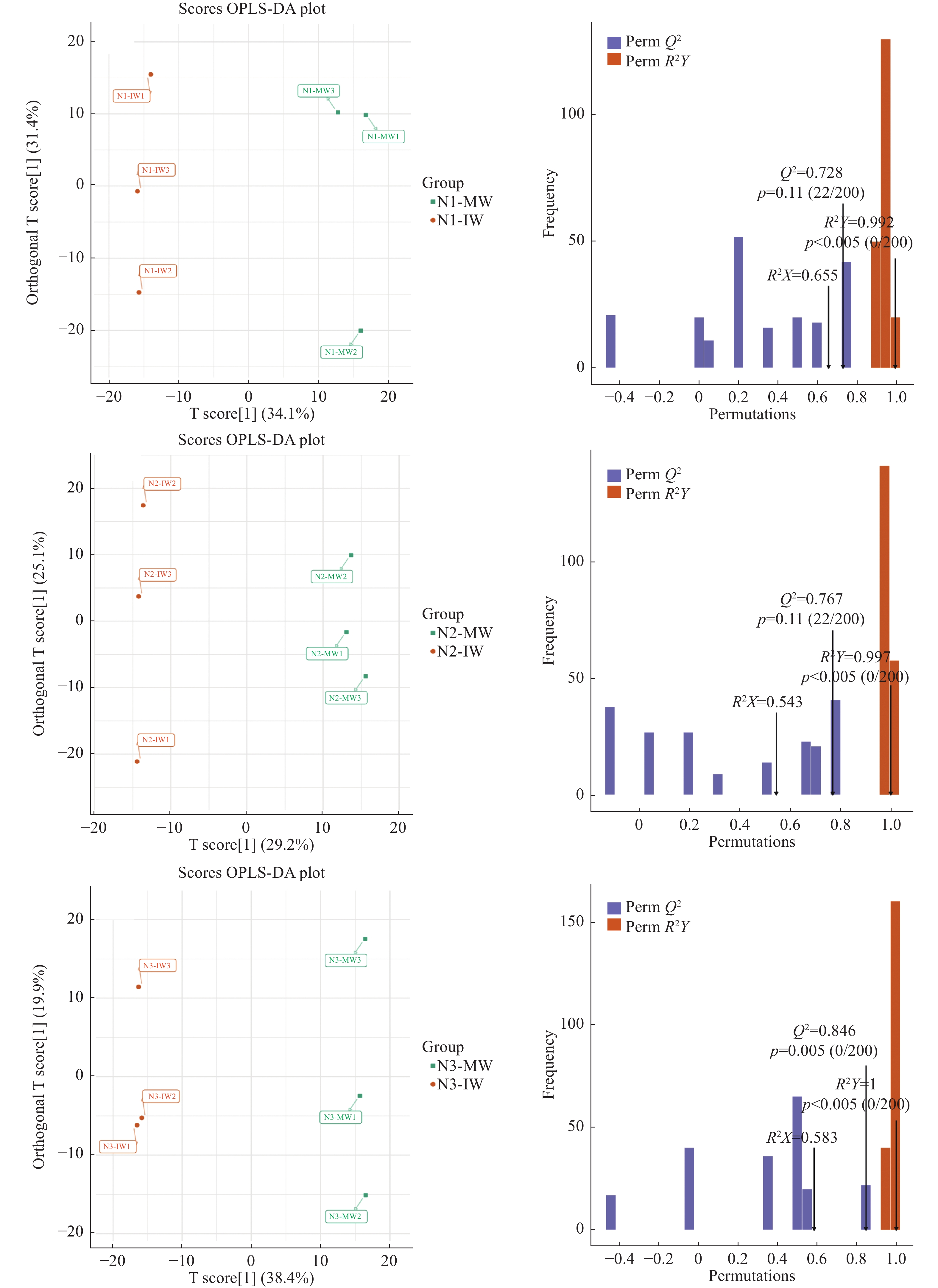
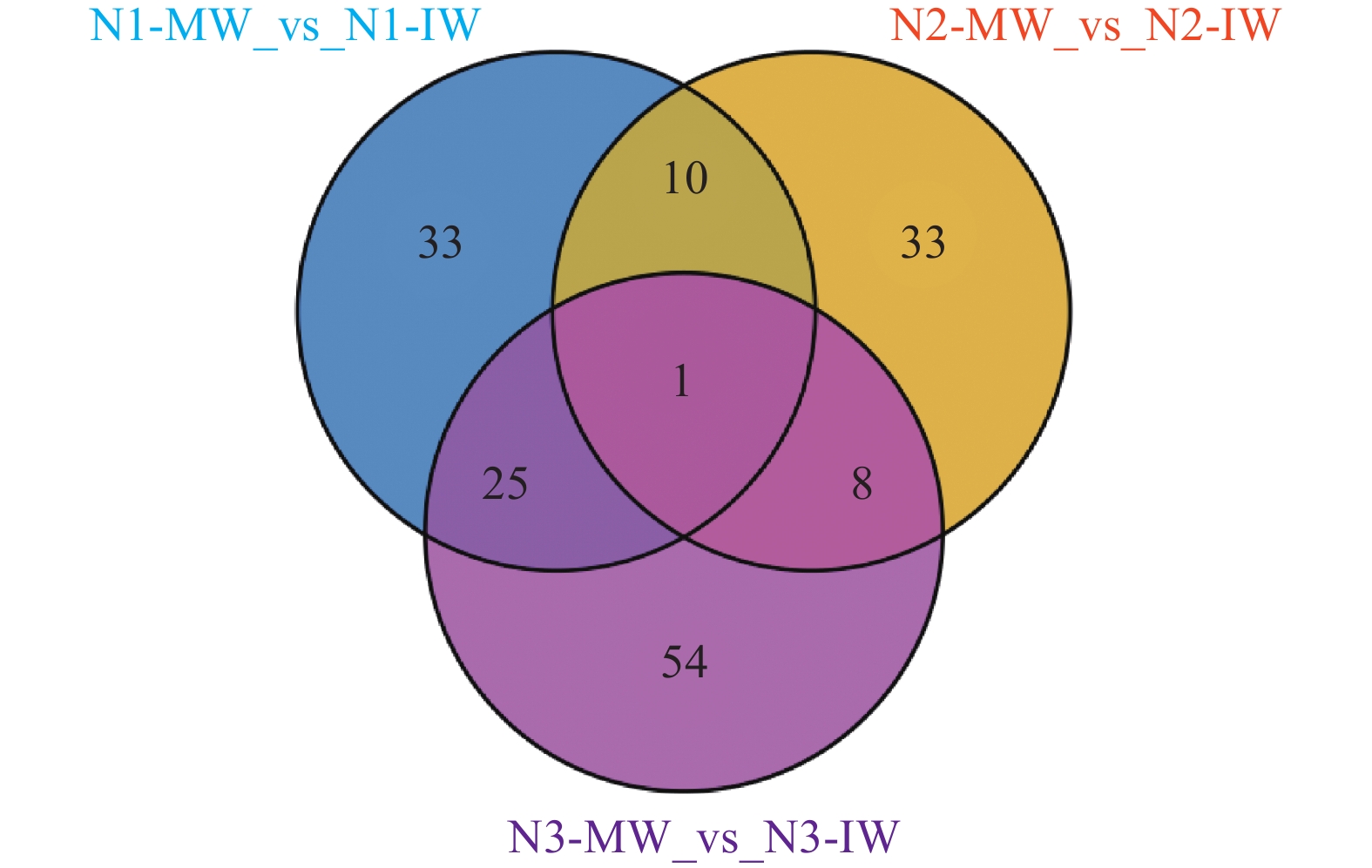
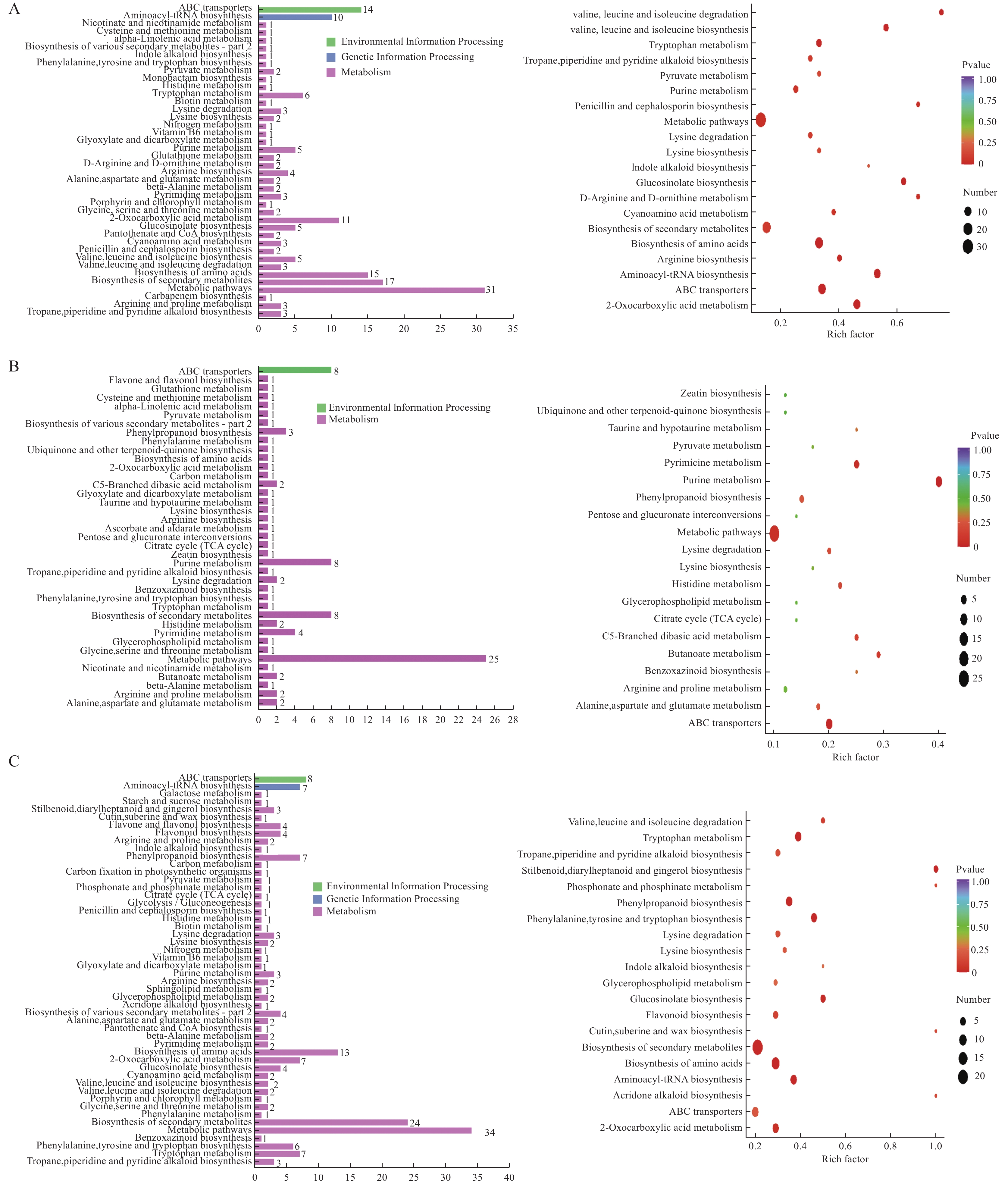
为了解单作、间作小麦响应白粉病侵染的代谢差异、揭示间作提高小麦抗白粉病生理机制, 本文通过盆栽试验设置 75 mg∙kg−1 (N1)、150 mg∙kg−1 (N2)、225 mg∙kg−1 (N3) 3个施氮水平, 研究接种白粉病病原菌后, 小麦单一种植和小麦蚕豆间作下白粉病的发病情况, 并通过广泛靶向代谢组学分析单作、间作小麦响应白粉病菌侵染的差异。结果表明: 氮水平和氮水平×种植模式显著影响小麦白粉病发病率和病情指数; 在3个氮水平下, 间作降低白粉病发病率25.54%~38.81%、降低病情指数20.11%~21.97%, 其中低氮水平控制效果较好。白粉病菌侵染后, 单作、间作小麦叶片中共检测到822种代谢产物, N1、N2和N3水平下分别发现差异代谢物69种、52种和88种。KEGG代谢通路分析发现单间作小麦差异代谢物主要富集在氨基酸的生物合成、代谢途径和次生代谢物的生物合成。其中N1和N2水平下, 差异代谢物富集在代谢途径, N1和N3水平下差异代谢物富集于氨基酸的生物合成。进一步对上调、下调差异倍数前10的代谢物分析发现, 与单作相比, N1水平间作上调了谷胱甘肽还原型、L-色氨酸、L-天冬酰胺和L-谷氨酰胺, N3水平间作上调了L-天冬酰胺、L-高甲硫氨酸和L-色氨酸。除此之外, 少数生物碱类、酚酸类和有机酸类等代谢物质在氮胁迫下也呈现不同程度的变化。总之, 单作和间作小麦响应白粉病病菌侵染的应答过程受氮水平调控。在白粉病病菌侵染中、间作调控差异代谢物如氨基酸及其衍生物类、生物碱类、酚酸类和有机酸类等在植物体内的变化, 可能是间作提高小麦白粉病抗性的机制之一。其中, 氮胁迫条件下间作调控氨基酸及其衍生物与小麦白粉病抗性提高密切相关。

洞悉有机农业发展的多元驱动路径对于促进我国农业绿色可持续发展至关重要。本文基于组态理论, 以中国省域有机农业发展为案例, 综合运用必要条件分析和模糊集定性比较分析方法, 通过探讨市场环境、资源禀赋、组织条件和政策制度的多重因素对区域有机农业发展的联动效应, 揭示有机农业发展的多元驱动路径。研究表明: 市场环境、资源禀赋、组织条件和政策制度等方面的多重因素并发联动形成了有机农业发展的多元条件组态; 存在4条驱动有机农业发展的差异化路径: 政策主导组织助力型、市场和组织双轮驱动型、认证和组织叠加带动型以及认证带动资源优势型。3种导致非高有机农业发展的条件组态均指向有机认证和产业化组织促进有机农业发展的显著效应。因此, 研究认为积极推动有机产品认证、发展产业化组织、加大政府正向激励是促进有机农业发展的主要政策建议, 此外各地应根据自身特点和资源禀赋选择有机农业发展的差异化路径。

微塑料作为一种新型环境污染物, 对生物体和自然环境的负面影响受到广泛关注, 而微塑料与重金属复合污染对于蔬菜作物影响的研究却少有报道。为探讨聚苯乙烯纳米塑料(PSNPs)、铅(Pb)及其复合污染对菠菜种子萌发和幼苗生长的影响, 研究了菠菜种子和幼苗分别暴露于200~1600 mg∙L−1 PSNPs、5~100 mg∙L−1 Pb及其复合溶液后发芽率、发芽势、发芽指数、根长、芽长, 超氧化物歧化酶(SOD)和过氧化物酶(POD)活性, 及可溶性蛋白含量的变化。结果表明, 单一的PSNPs (≥400 mg∙L−1)胁迫会显著降低种子的发芽率、发芽势和发芽指数, 低浓度的PSNPs (200 mg∙L−1)胁迫显著促进菠菜种子根、芽的伸长, 高浓度(1600 mg∙L−1)的PSNPs胁迫显著抑制SOD、POD的活性, 不同浓度的PSNPs均会增加可溶性蛋白的含量, 但仅在800 mg∙L−1浓度组显著高于对照组。 单一的Pb (≥25 mg∙L−1)胁迫抑制菠菜种子的萌发, 降低SOD的活性, 而提高POD活性和可溶性蛋白的含量。PSNPs和Pb的复合污染表明, 相比于Pb单一胁迫, PSNPs与Pb复合污染对菠菜种子的萌发起拮抗作用, 降低了Pb单独胁迫对种子萌发的抑制作用; 而PSNPs-Pb复合污染对菠菜幼苗的影响主要是低浓度(200 mg∙L−1)PSNPs与Pb二者表现为协同作用, 高浓度(800 mg∙L−1) PSNPs与Pb复合污染加重了对菠菜幼苗的毒害, 主要表现为SOD和POD活性的显著降低。研究表明, PSNPs能够缓解Pb对菠菜种子萌发的抑制作用; 低浓度PSNPs (200 mg∙L−1)与Pb对菠菜幼苗的影响表现为协同作用, 而高浓度(800 mg∙L−1)PSNPs与Pb对菠菜幼苗主要表现为拮抗作用。
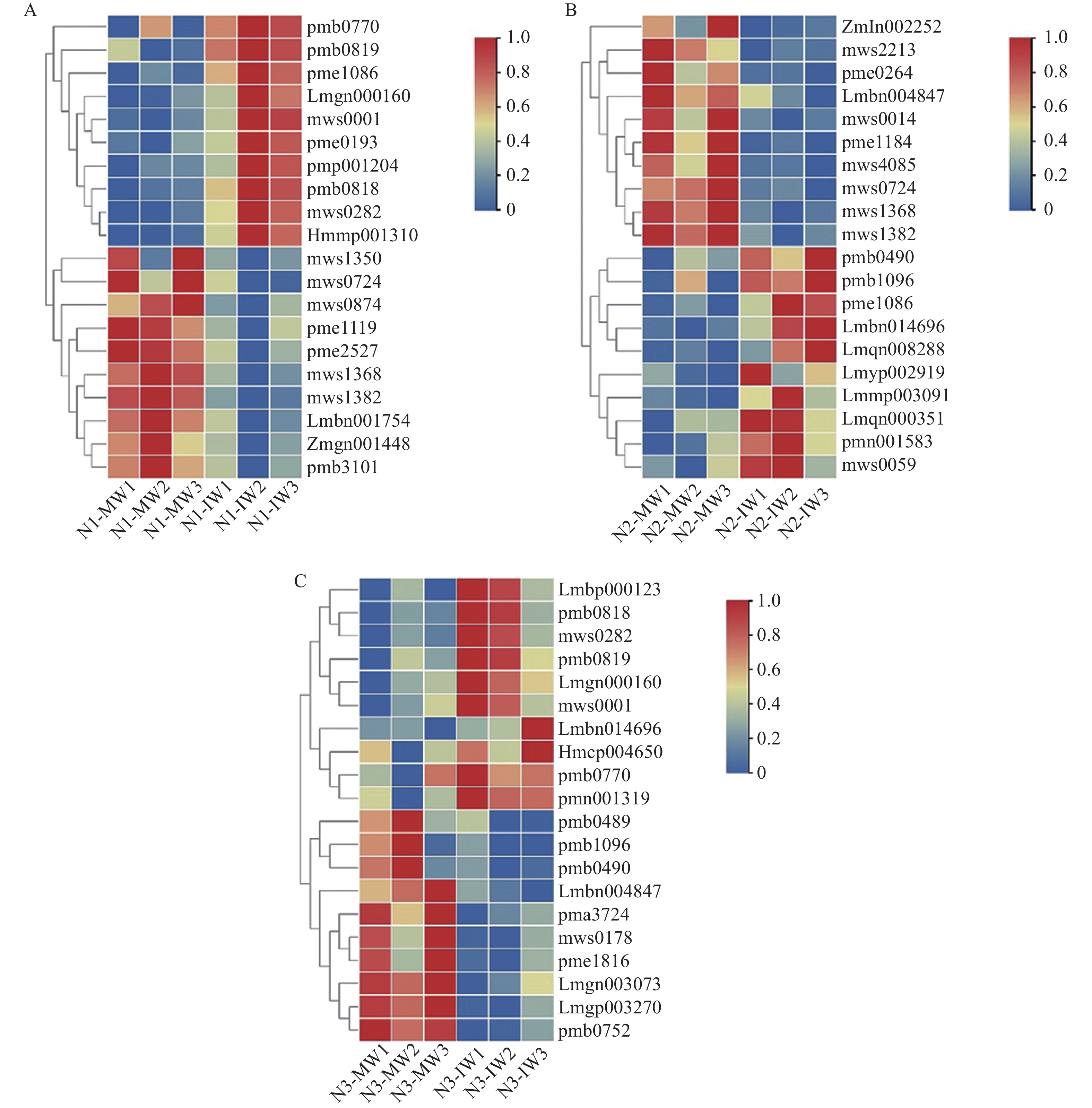
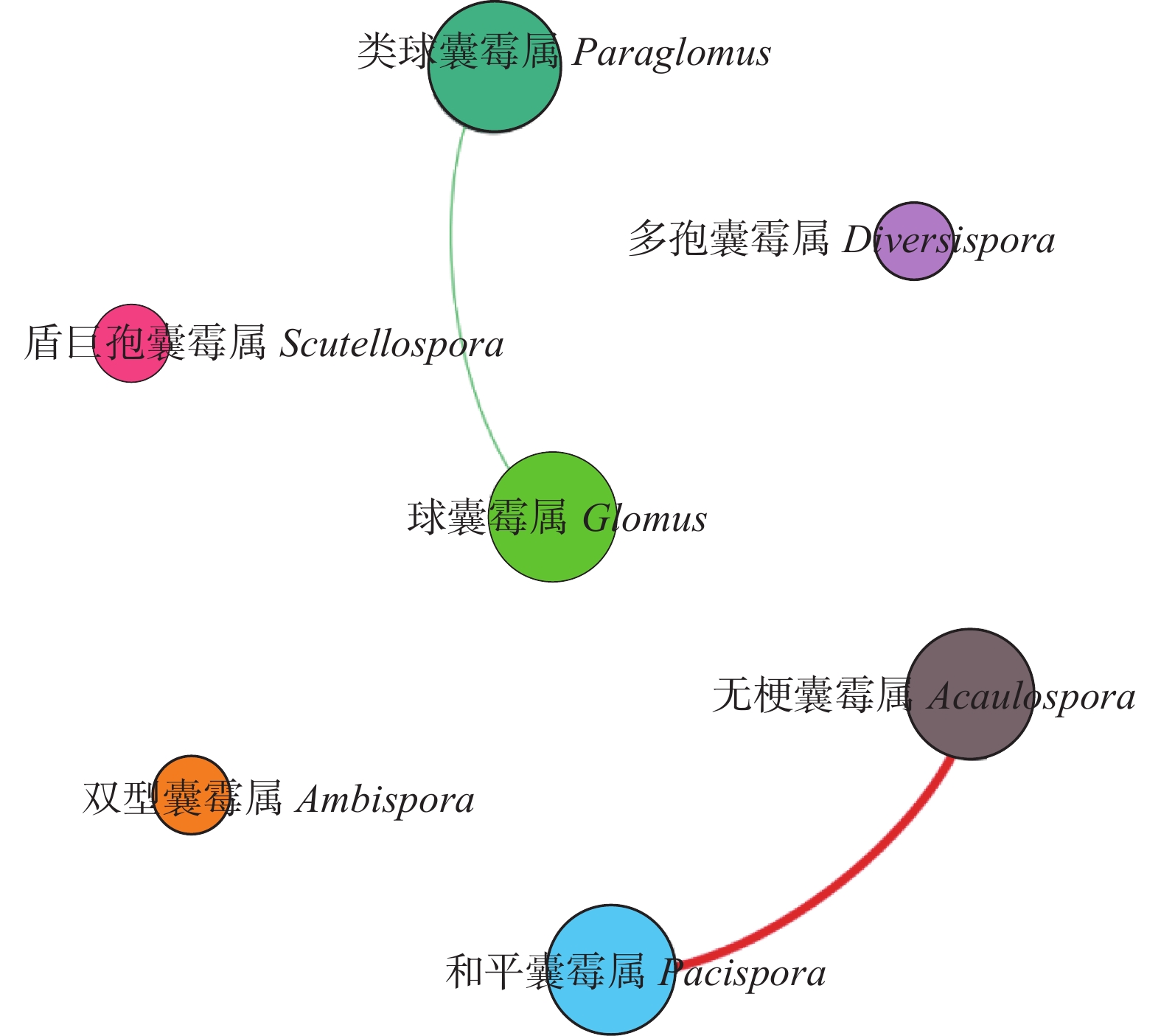
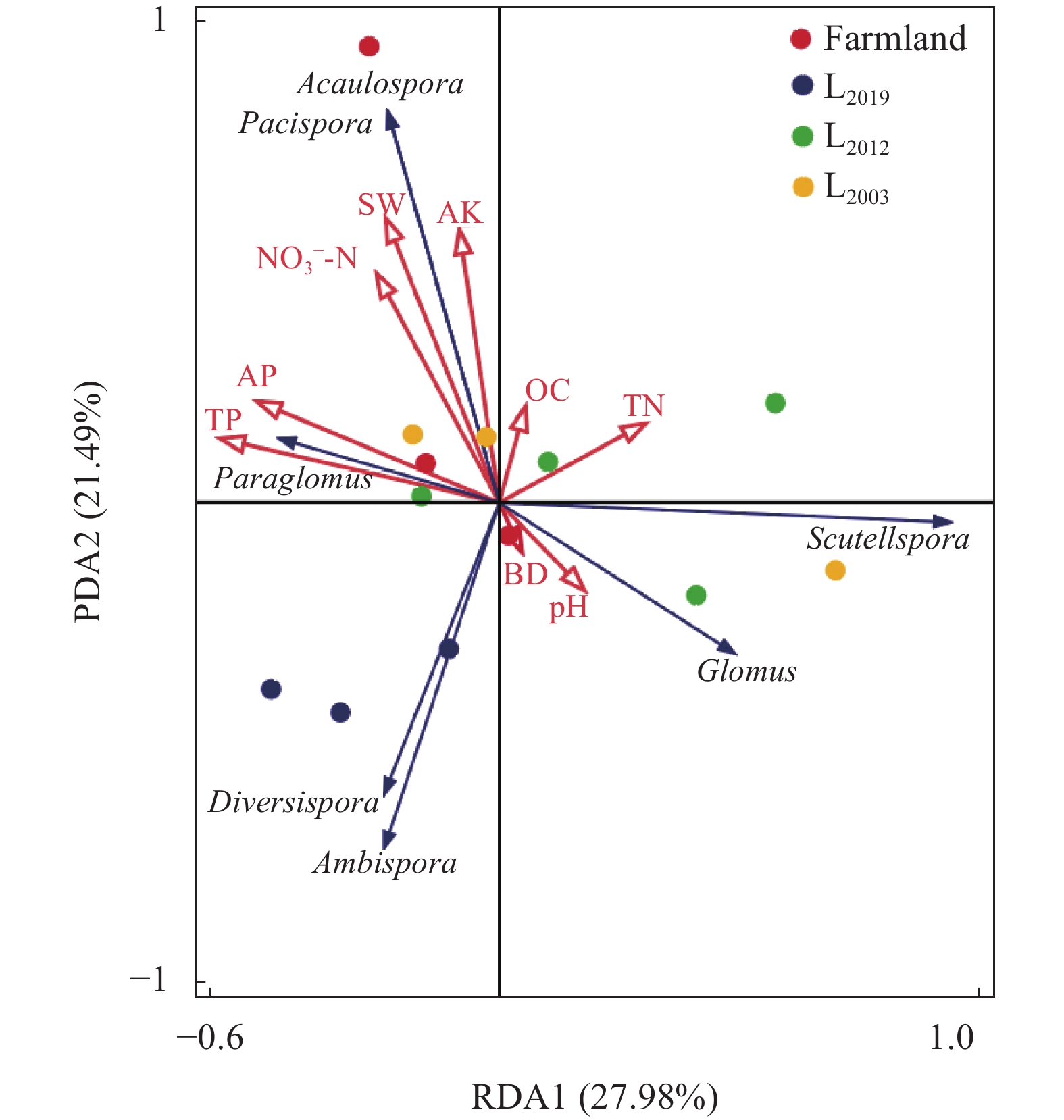
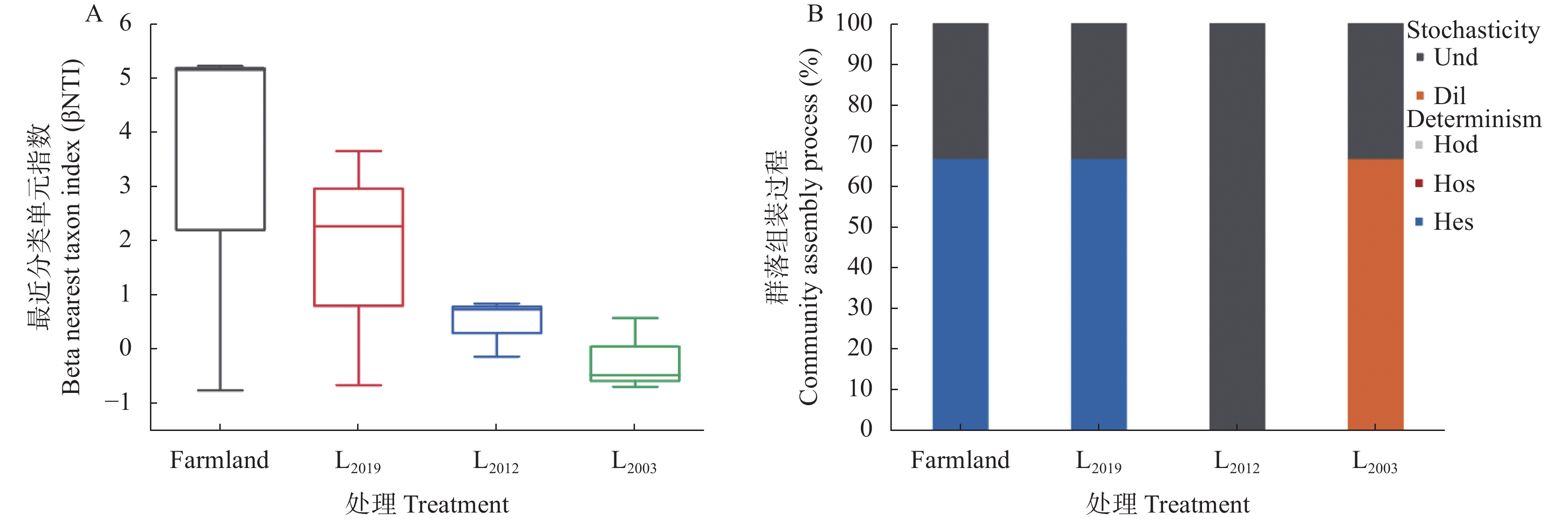
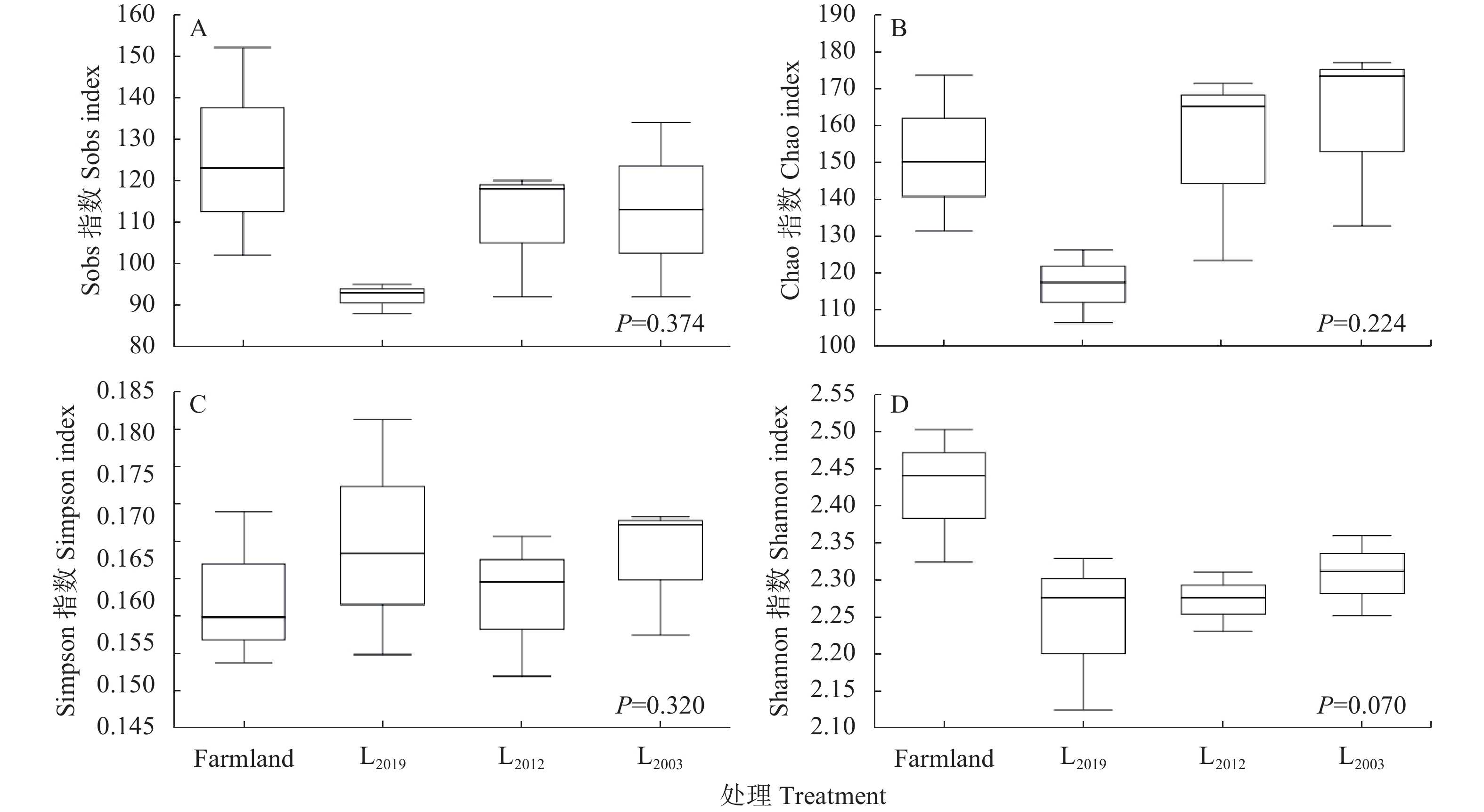
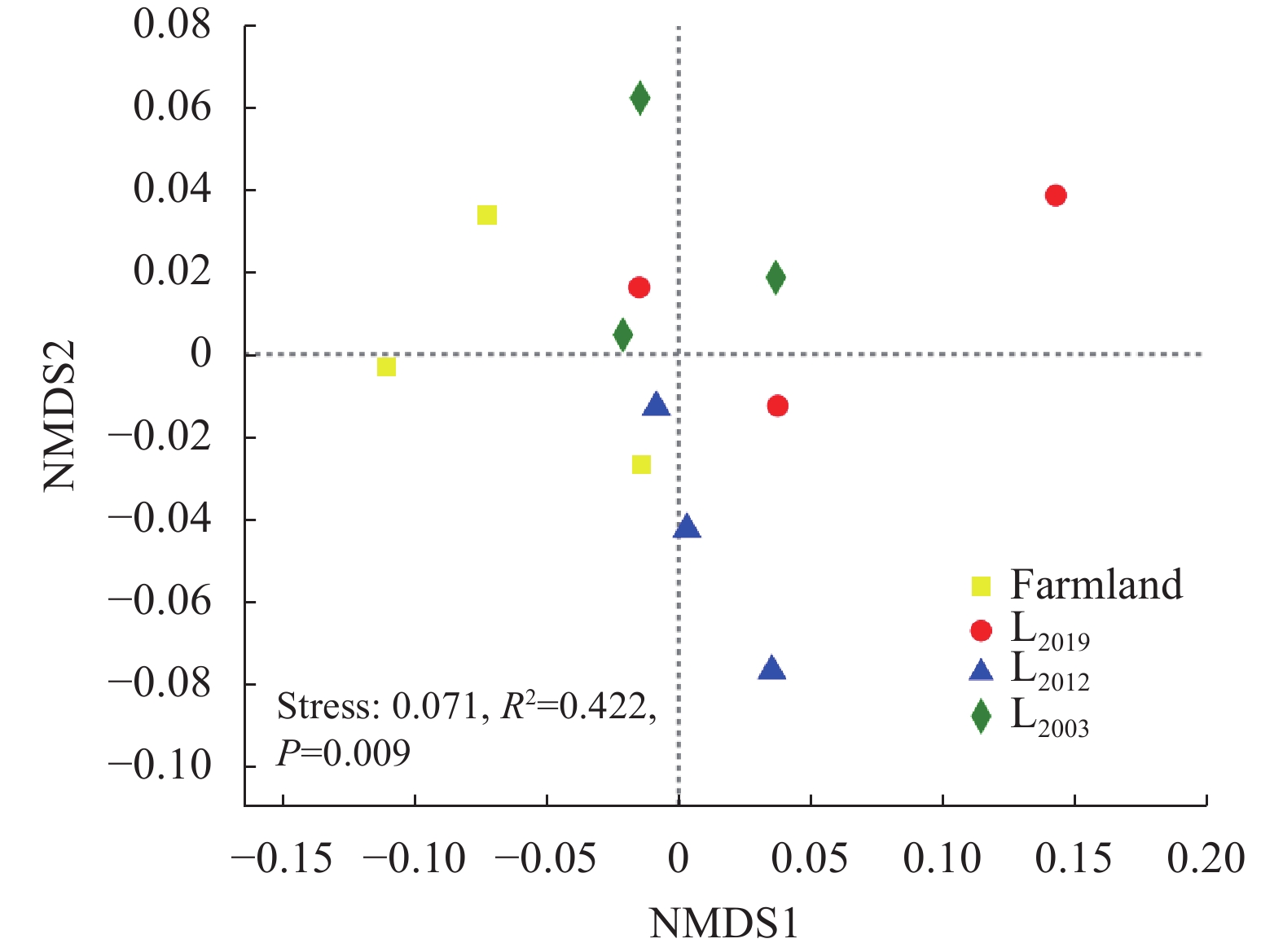
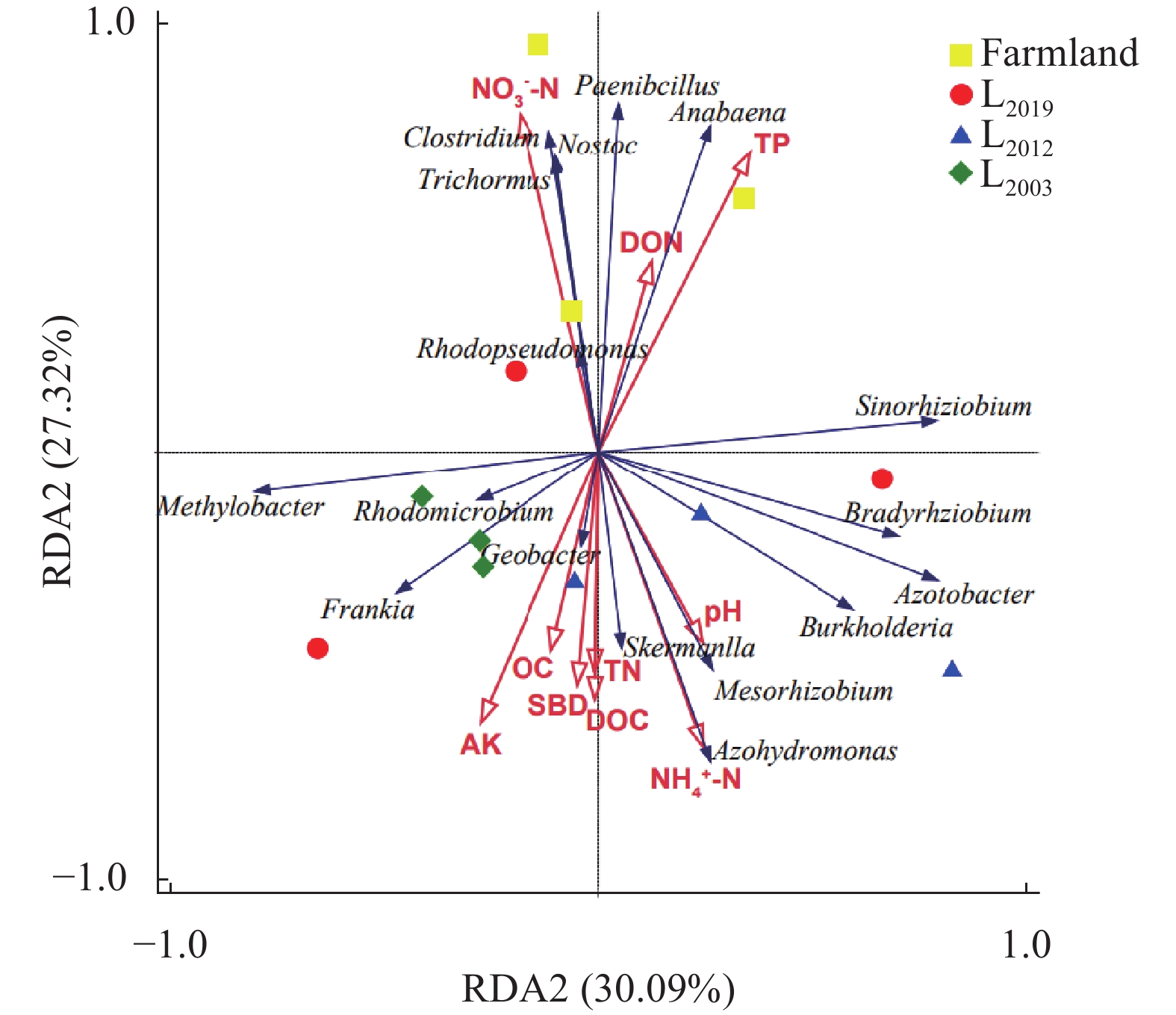
为揭示多年种植苜蓿对土壤丛枝菌根真菌(AMF)群落结构和多样性的影响, 本研究通过布设在黄土高原半干旱区的田间试验, 基于2019、2012和2003年建植的紫花苜蓿(Medicago sativa), 以农田玉米(Zea mays)为对照, 采用高通量测序和PCR技术, 结合分子生态网络研究不同种植年限紫花苜蓿地土壤AMF群落组成和丰度, 并基于零模型揭示了土壤AMF群落的组装过程。结果表明: 黄绵土区AMF属于球囊菌门的1纲4目7科7属, 球囊霉属、类球囊霉属和多孢囊霉属为紫花苜蓿地和农田土壤共有类群, 且均以球囊霉属(65.15%~99.12%)为优势属, 其主要贡献了不同处理分组中土壤AMF群落结构的改变。长期种植紫花苜蓿使得和平囊霉属和无梗囊霉属消亡, 但促生了双型囊霉属和盾巨孢囊霉属, 其中双型囊霉属相对丰度表现为L2019处理显著高于其他处理(P<0.05)。网络关联分析发现, 高丰度的球囊霉属和类球囊霉属之间呈现负相关, 而低丰度的和平囊霉属和无梗囊霉之间呈现正相关。基于零模型的群落组装结果表明, 农田与L2019处理由确定性过程主导(66.67%), L2012和L2003处理由随机性过程主导(100%), 这表明长期种植紫花苜蓿形成稳定的土壤环境使其随机性过程增加, 利于维持人工草地生态系统功能的可持续性和稳定性。


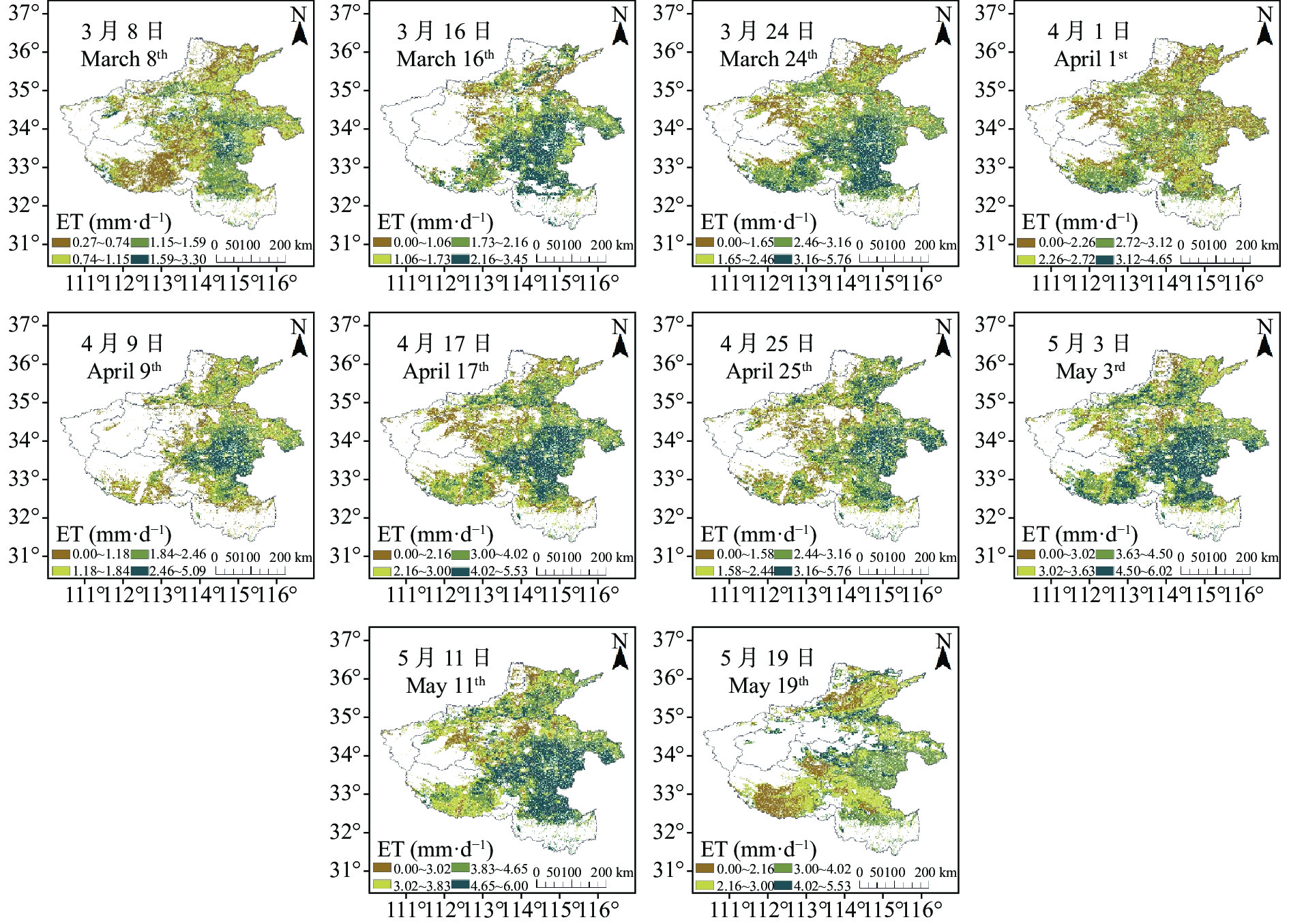
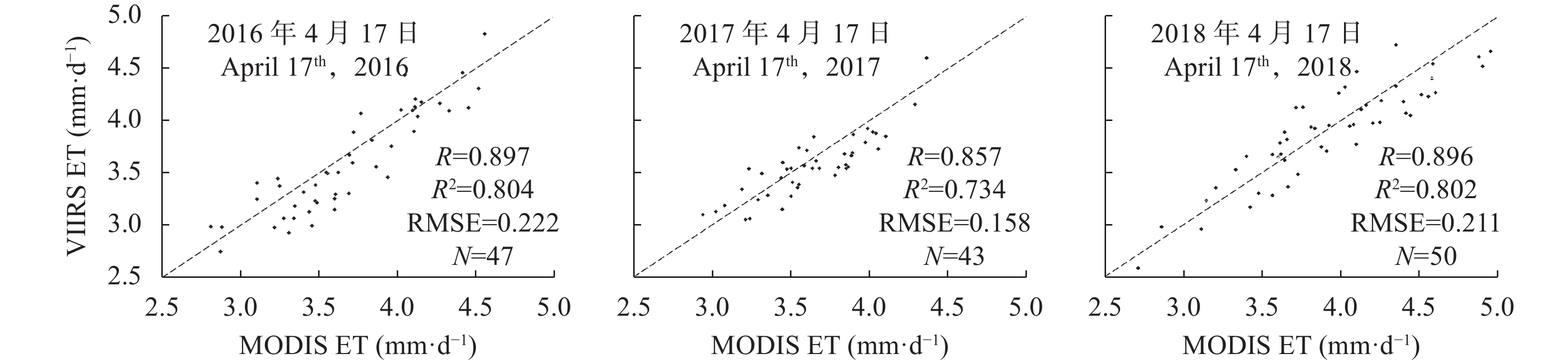
农田蒸散量 (evapotranspiration, ET)是农田水热交换过程的关键变量, 准确估算农田ET对了解农田土壤水分变化动态, 监测预测作物旱情, 指导科学灌溉等具有重要作用。将Suomi NPP (National Polar-orbiting Partnership)卫星的新型遥感数据源可见光红外成像辐射仪(Visible Infrared Imaging Radiometer Suite, VIIRS)数据用于ET反演, 基于地表能量平衡理论, 将NPP VIIRS反演的地表温度、地表反照率等参数和优化计算的VIIRS NDVI (normalized difference vegetation index)数据, 与SRTM DEM数据和气象观测数据输入地表能量平衡系统(surface energy balance system, SEBS)模型, 估算了2016—2018年河南省冬小麦关键生育时期——返青期至灌浆期的农田ET (VIIRS ET)。分别对比了VIIRS ET与彭曼公式(Penman-Monteith, P-M)计算的P-M ET、大型土壤蒸渗仪实测Real ET和MODIS (moderate-resolution imaging spectroradiometer)数据估算ET (MODIS ET)的结果, 并进行了ET时空变化特征分析。结果表明, VIIRS ET与Real ET的RMSE为0.203 mm∙d−1, 且VIIRS ET与P-M ET和MODIS ET均具有较高的一致性。该方法可为利用NPP VIIRS数据和SEBS模型估算ET提供方法、技术上的支持。研究区冬小麦关键生育时期ET的时间特征表现为自返青期后日逐渐上升, 抽穗期达到最大值, 灌浆期开始下降的趋势, 与冬小麦生长发育规律具有较好的一致性。综合空间特征来看, 在关键生育时期河南省冬小麦ET均呈现中部和东南部较高, 向西北部和西南部逐渐降低的趋势, 与灌溉条件具有较强的对应性。河南省冬小麦ET时空特征可为河南省农业水资源管理、分配和高效利用等提供重要依据。
为了探究安全和环境友好的烟草黑胫病的绿色防控方式, 本研究用‘地隆1号’芥菜采用培养皿培养方法进行生物熏蒸, 研究了其对烟草疫霉菌(Phytophthora nicotianae)的抑制作用。采用菌丝生长速率法测定了其对烟草疫霉菌(A2 交配 型)Pp025菌丝生长的抑制情况, 对烟草疫霉菌孢子囊形成及游动孢子释放、孢子萌发及芽管伸长和卵孢子形成的影响, 通过电导率和丙二醛含量变化测定了其对烟草疫霉菌细胞膜透性的影响; 采用离体叶片法测定了芥菜熏蒸处理后烟草疫霉菌致病力的变化。结果显示: 利用‘地隆1号’芥菜进行生物熏蒸可显著抑制烟草疫霉菌的菌丝生长, 其半最大效应浓度(EC50)值为0.360 g, 75%最大效应浓度(EC75)值为0.499 g, 最小抑菌量(MIC)为0.500 g, 抑制效果与‘地隆1号’芥菜鲜物质的量呈剂量效应。0.300 g芥菜鲜物质即可显著抑制孢子囊的形成及游动孢子的释放, 0.500 g芥菜鲜物质熏蒸处理对Pp025孢子囊形成和游动孢子释放的抑制率分别达72.8%和86.6%; 当芥菜鲜物质的量增加为0.700 g时, 可完全抑制Pp025孢子囊的形成; 0.300 g和0.400 g鲜物质即可分别完全抑制游动孢子萌发及芽管伸长。‘地隆1号’芥菜鲜物质熏蒸处理可减少Pp025卵孢子的形成, 且减少幅度与芥菜鲜物质的量呈剂量效应, 0.700 g鲜物质熏蒸处理可完全抑制卵孢子的形成。同时, 熏蒸处理后增大了菌丝体细胞膜透性并导致膜脂质过氧化, 经熏蒸处理150 min后, Pp025的电导率为对照组的2.2倍, 丙二醛含量是对照的8.0倍; 熏蒸处理减弱了病菌的致病力, 与对照相比, 病斑减小面积达88.9%。本研究结果表明: ‘地隆1号’芥菜鲜物质熏蒸处理对烟草疫霉菌具有很好的抑制和杀灭作用, 且熏蒸效果与‘地隆1号’芥菜鲜物质的量呈剂量效应。研究结果对生产上利用‘地隆1号’芥菜防控烟草黑胫病提供了理论依据。





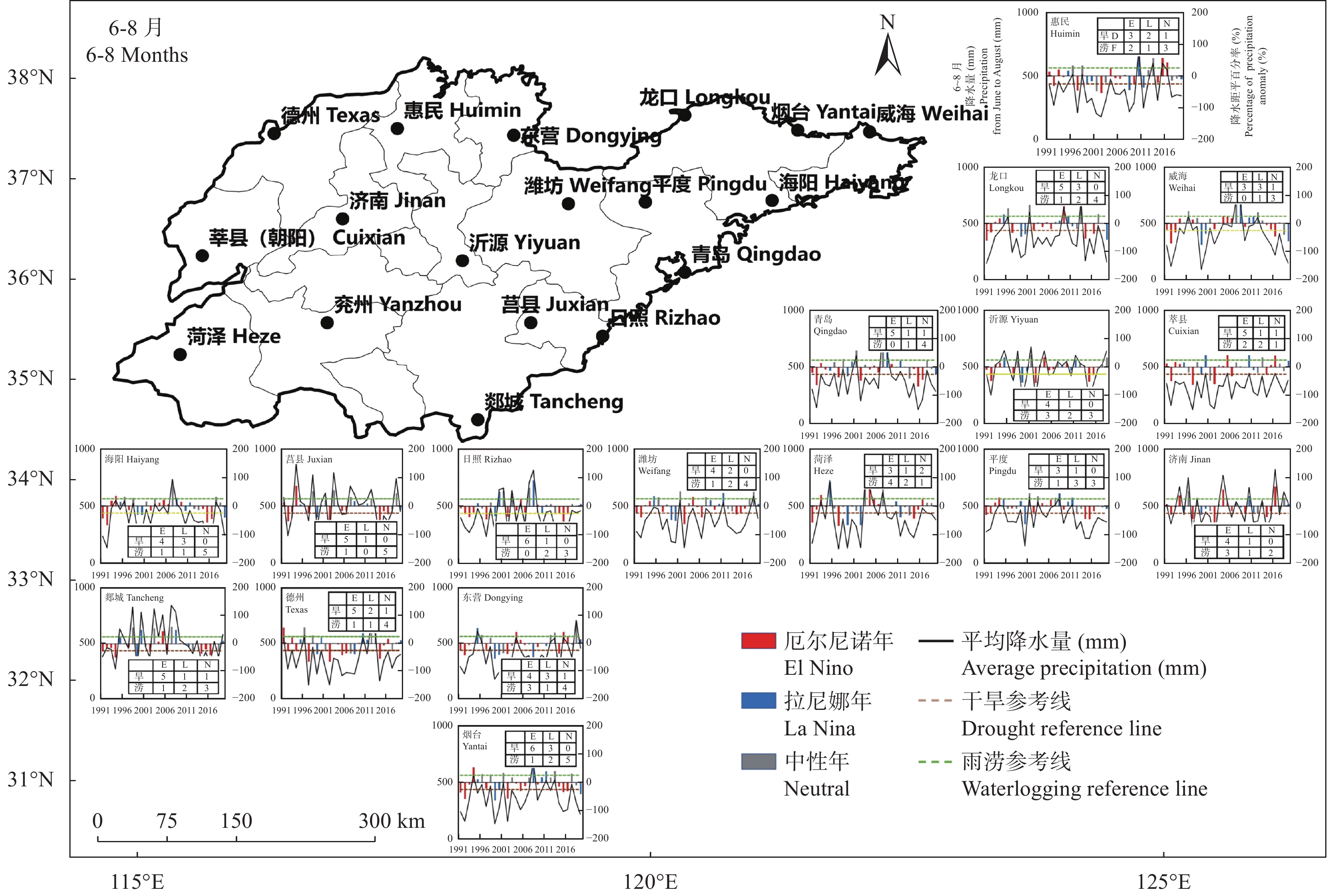
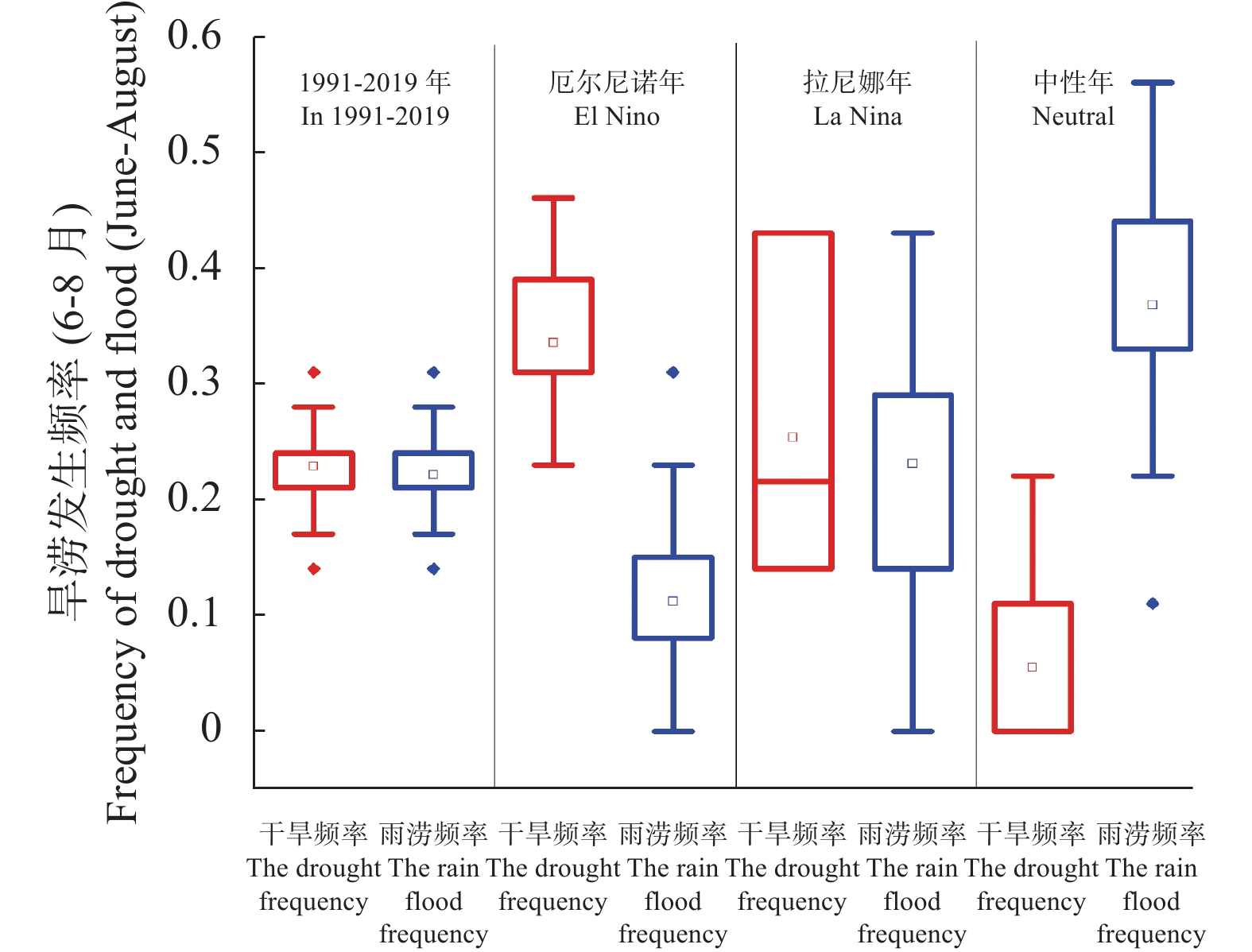
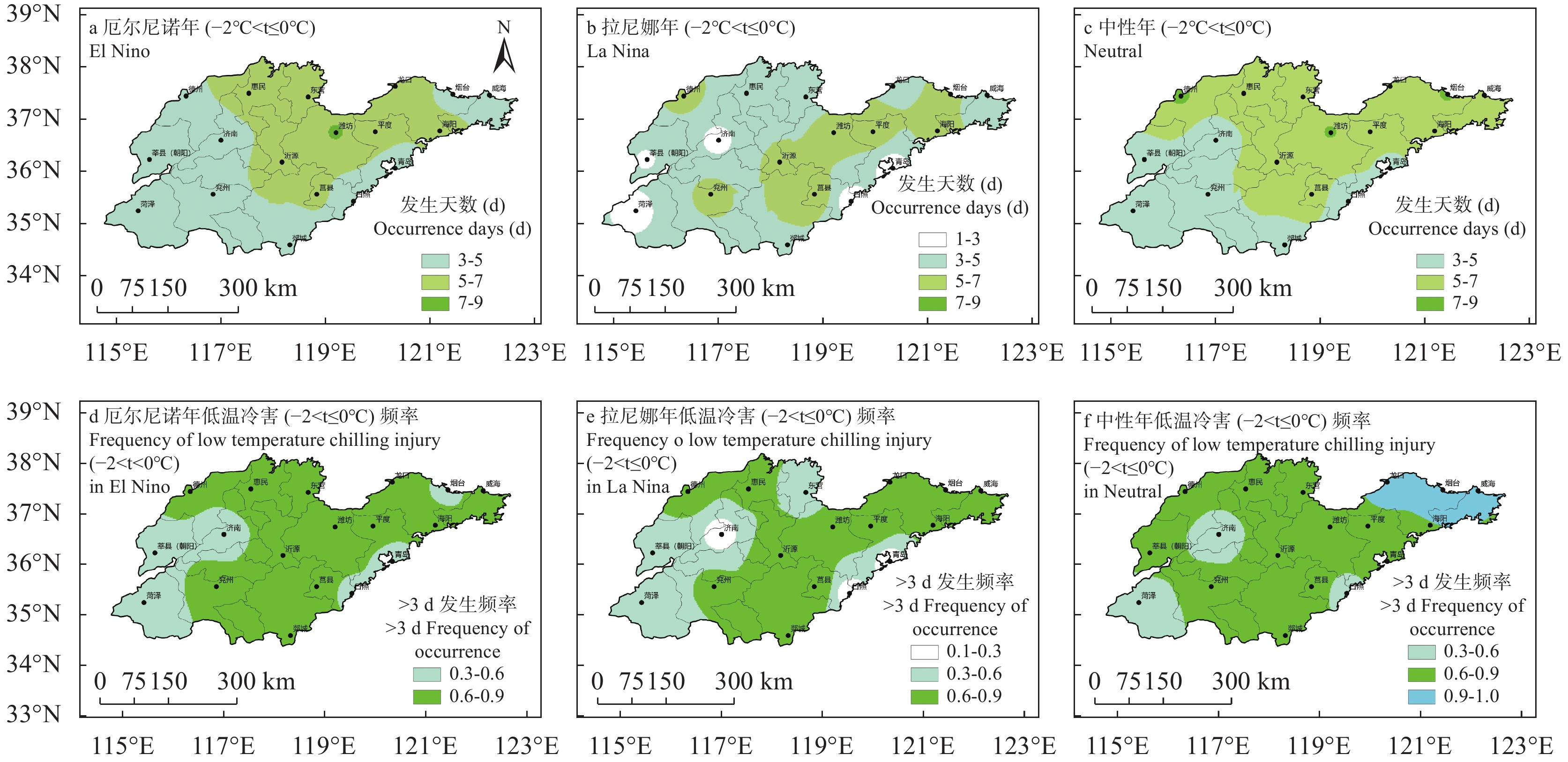
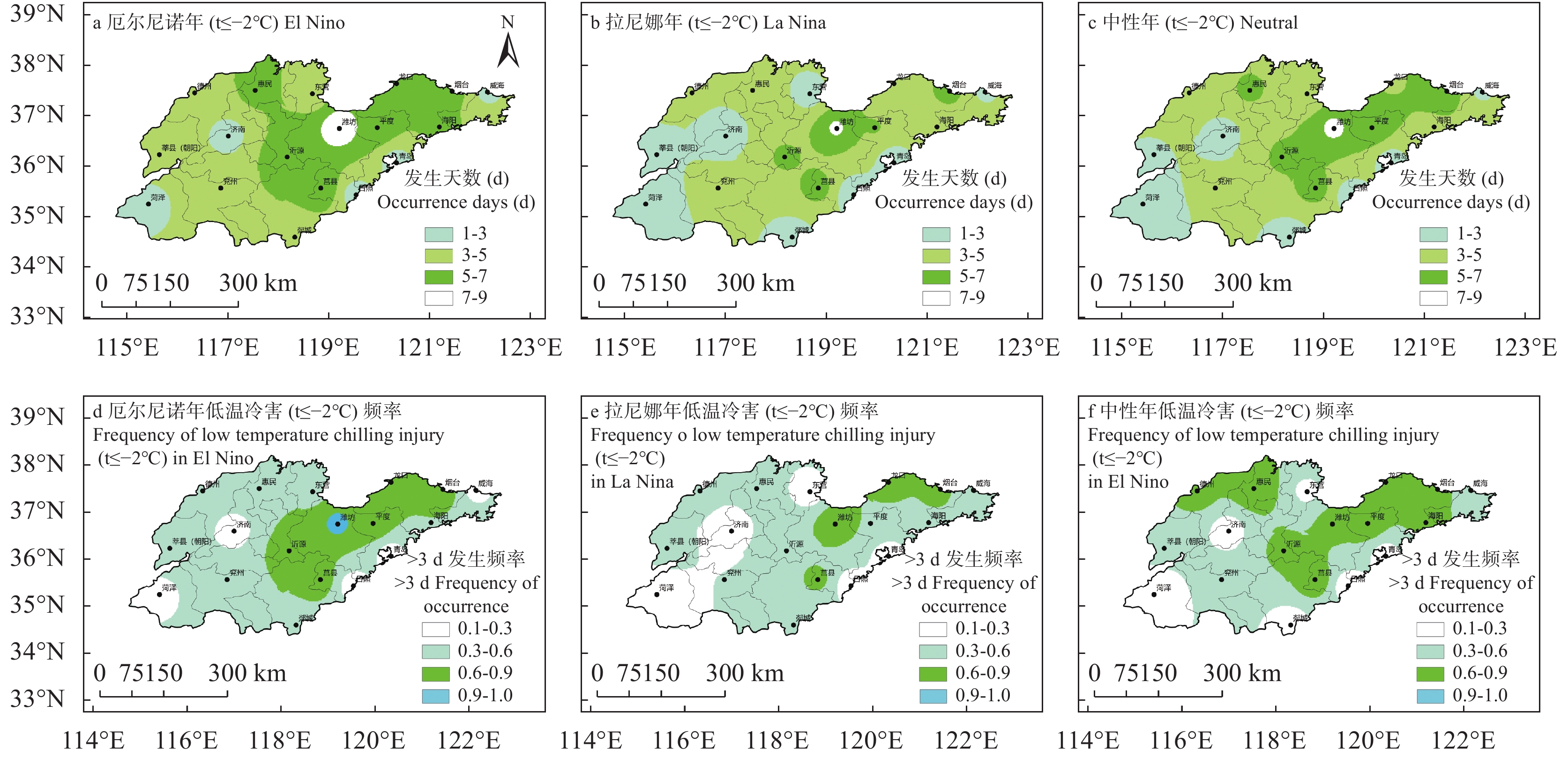
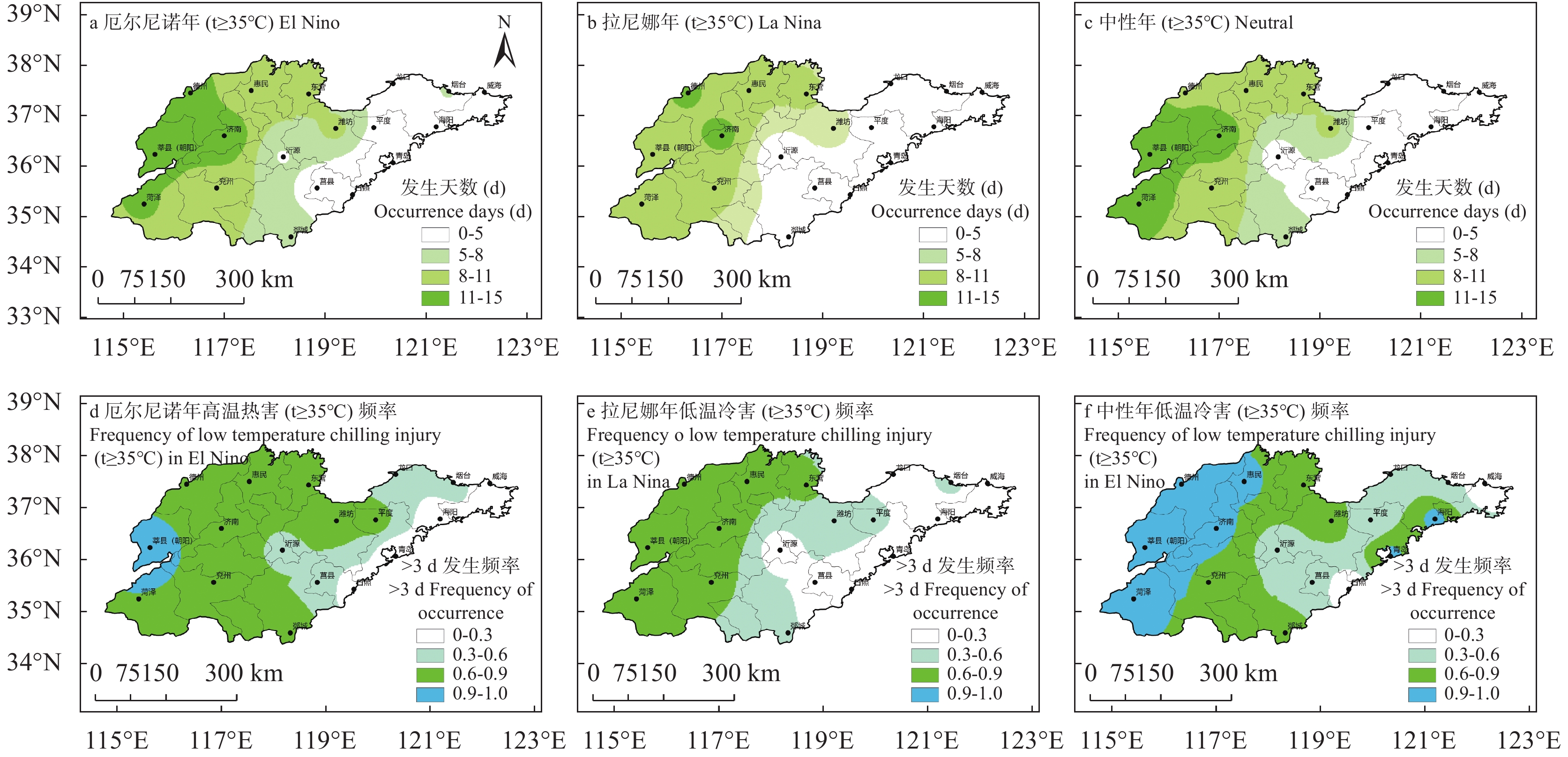
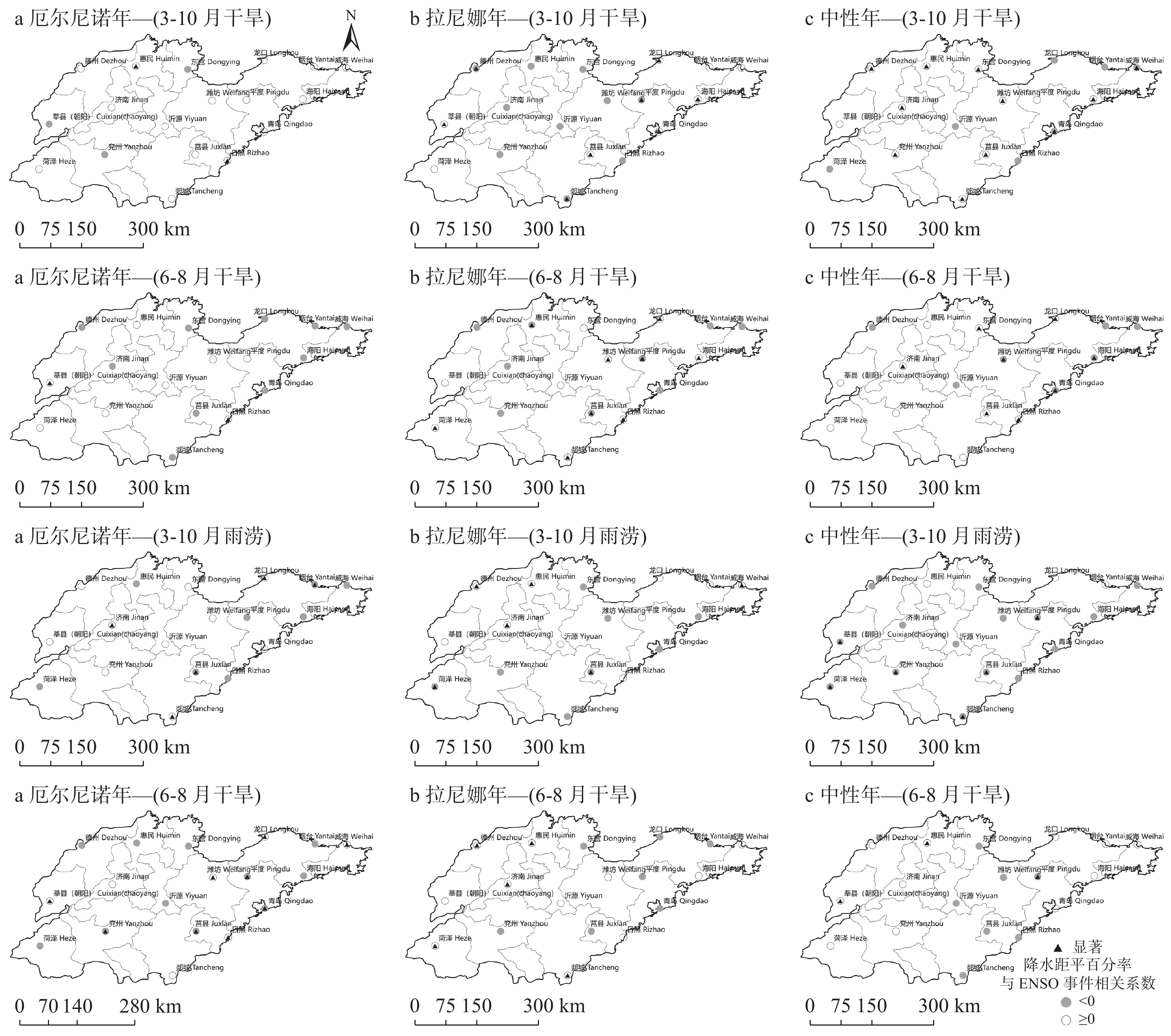
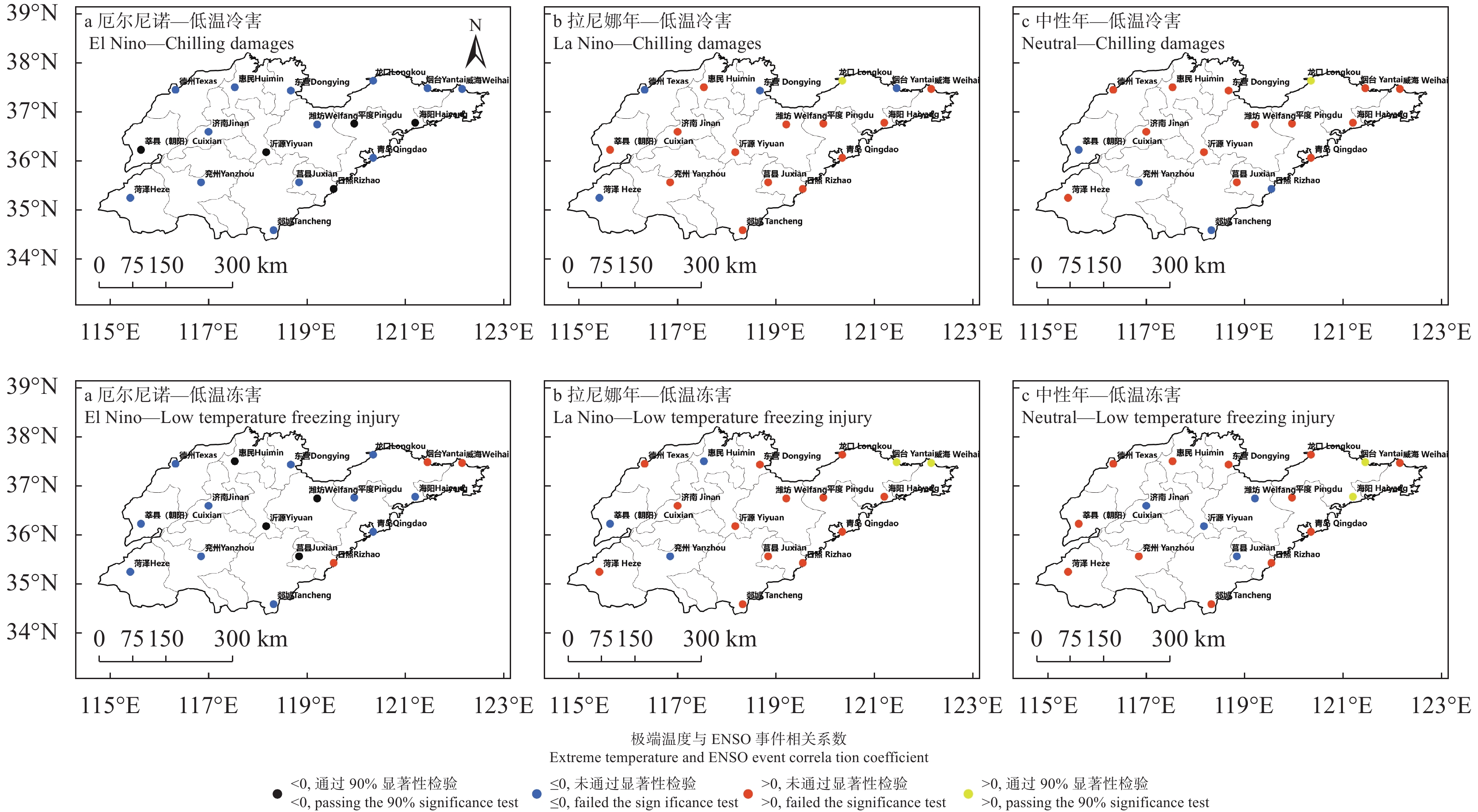
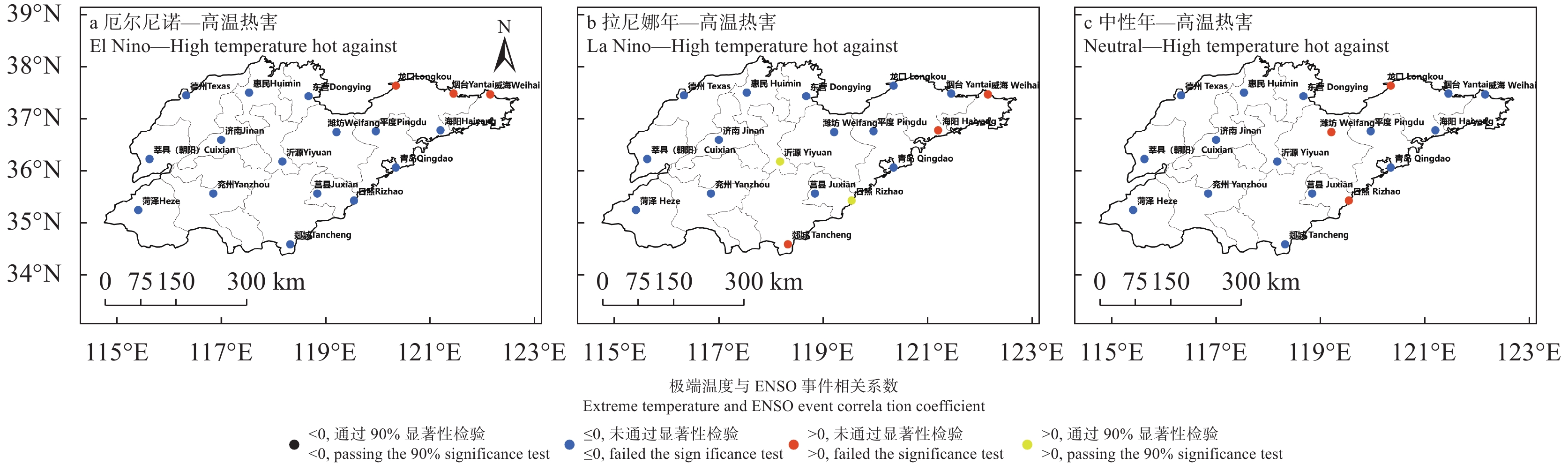
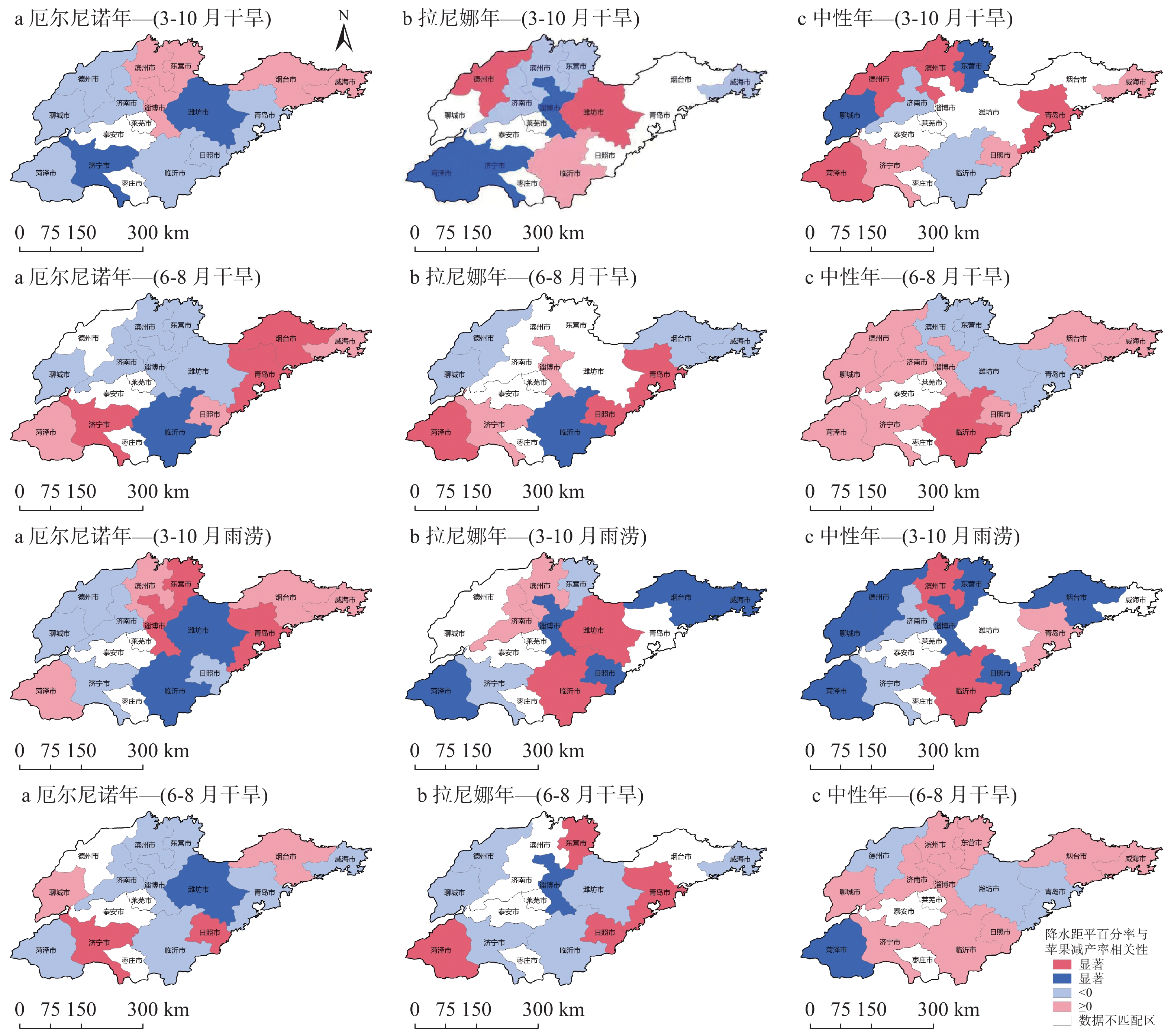
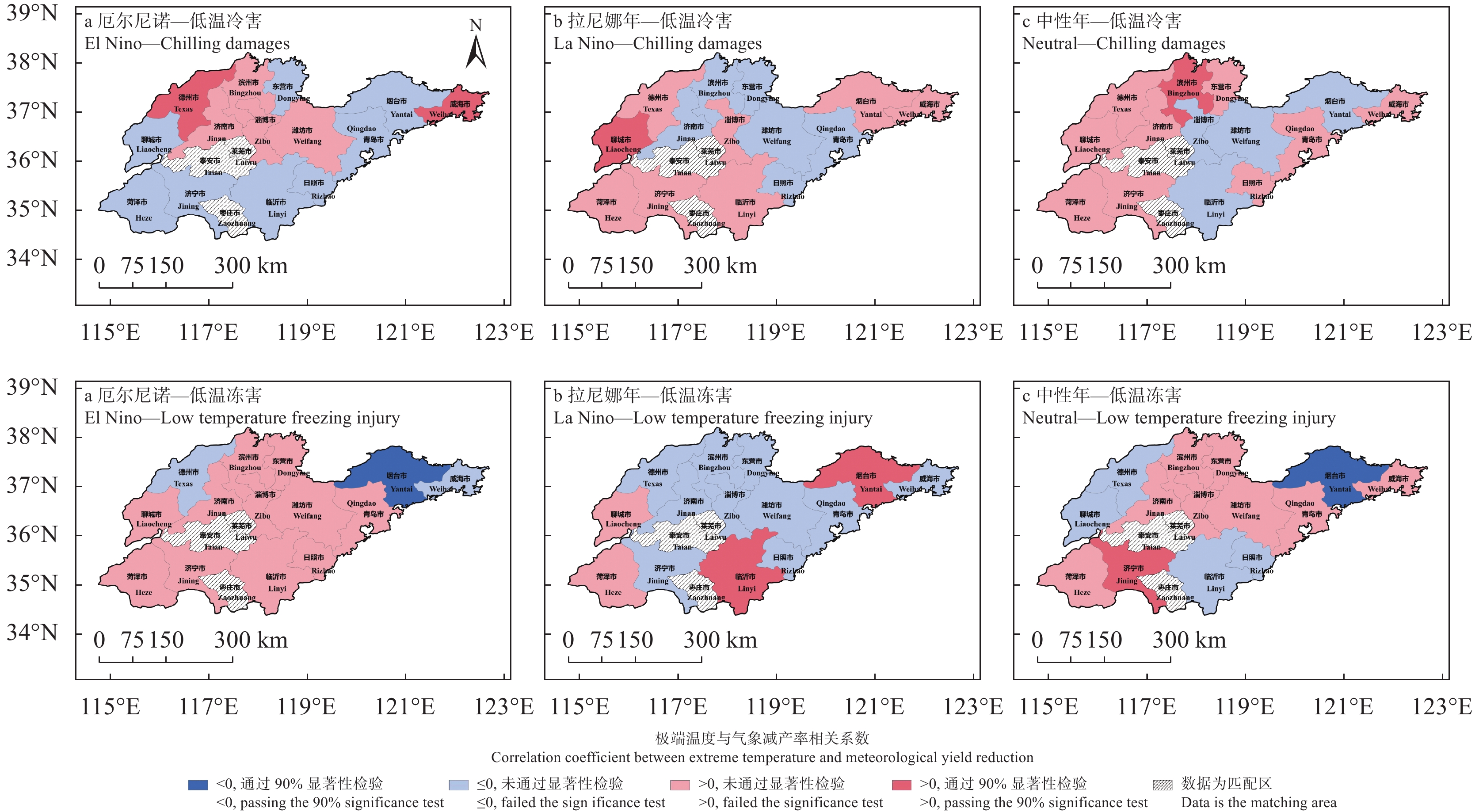
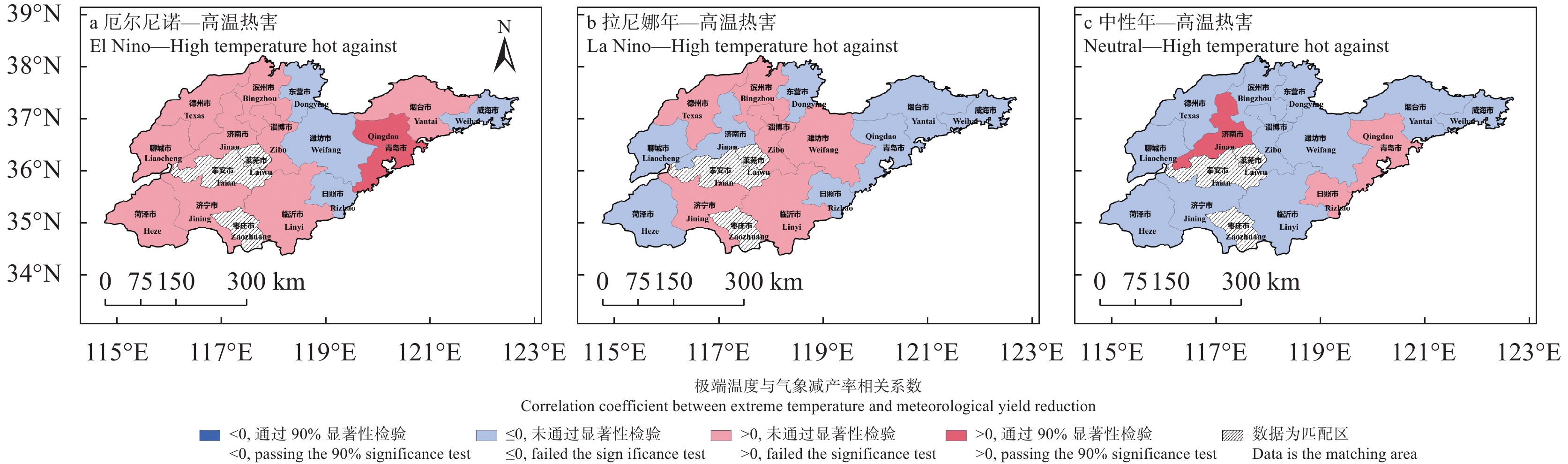
苹果作为山东优势果品之一, 其生产受农业气象灾害影响较大。探究在ENSO事件下山东农业气象灾害演变规律及其对山东苹果产量的影响, 对指导当地苹果生产具有重大意义。本文基于山东1991—2019年逐日气象观测数据、地市级苹果种植统计数据及ENSO事件数据, 利用数理统计分析和ArcGIS空间表达, 得出以下结论1) 1991−2019年不同ESNO年型下农业气象灾害发生情况区域差异显著。6—8月果实膨大期厄尔尼诺年干旱灾害发生较为频繁, 共计78次, 干旱频率最高约50%; 中性年雨涝灾害较为严重, 高达60次。鲁西、鲁中等热量资源充足地区, 干旱发生较为频繁; 鲁南降水资源较为充沛地区, 雨涝灾害发生频繁。鲁东、胶东半岛等地3—5月苹果花期极端低温灾害发生较为频繁, 发生日数约7~9 d·a−1, 频率约为60%—100%。鲁西等地是6—8月苹果果实膨大期高温热害的高发区, 发生天数11~15 d·a−1。2)不同ESNO年型下, 干旱与厄尔尼诺年呈正相关, 与拉尼娜年呈负相关。3—10月苹果可生长期厄尔尼诺年南方涛动指数与雨涝呈正相关相关, 拉尼娜年、中性年南方涛动指数与雨涝呈负相关。3—5月苹果花期低温灾害与厄尔尼诺年南方涛动指数呈负相关; 与拉尼娜年、中性年南方涛动指数呈正相关。3) 3—10月苹果可生长期, 厄尔尼诺年, 胶东半岛地区干旱加剧, 导致苹果减产率上升; 中性年, 雨涝灾害, 也使得苹果减产减收影响加重。6—8月苹果果实膨大期, 拉尼娜年、中性年下, 鲁西地区干旱与苹果减产率呈正相关; 中性年, 山东大部分地区雨涝与苹果减产率呈正相关。厄尔尼诺年苹果减产率受极端低温灾害影响较小, 高温热害影响较大; 拉尼娜年、中性年山东大部分地区低温冷害、冻害天数增加, 导致苹果减产率上升, 风险加大。苹果生产中, 谨防厄尔尼诺年高温、干旱, 拉尼娜年、中性年应预防低温、雨涝灾害对苹果产量、品质的损害, 确保苹果产业而健康可持续的生产。

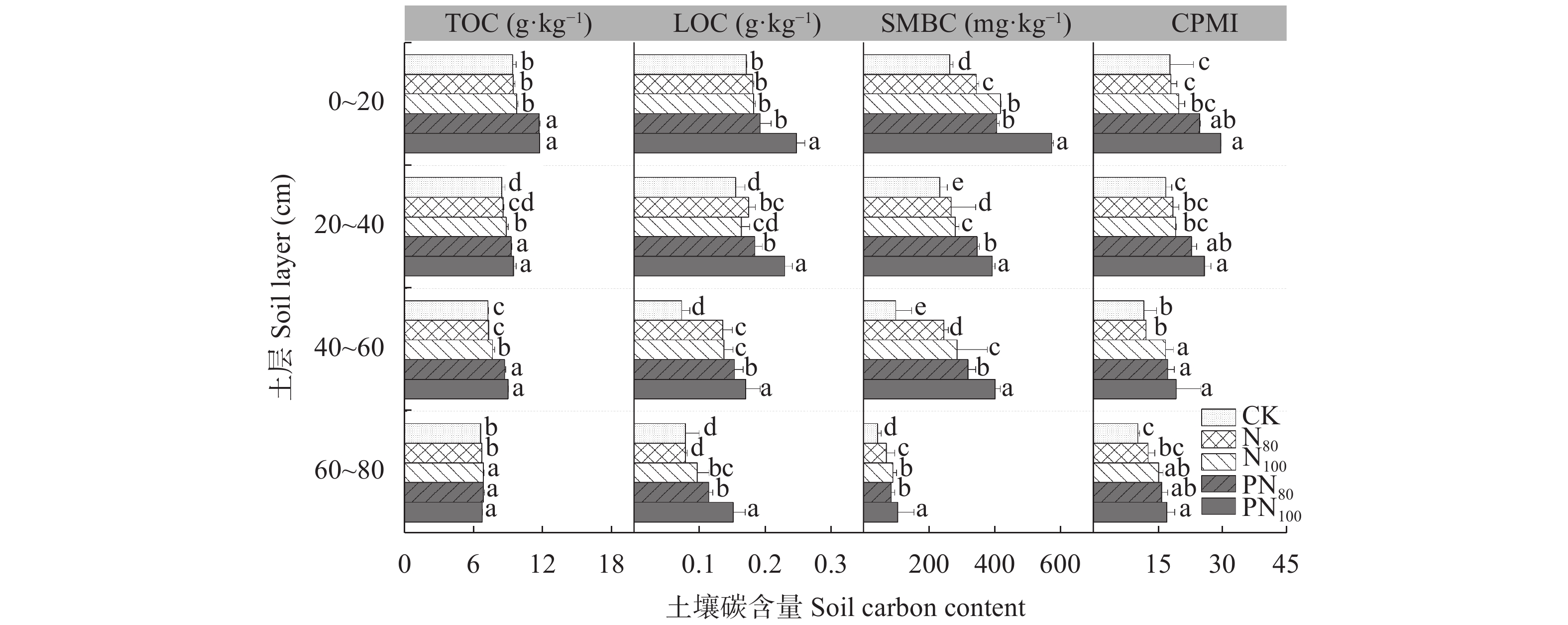
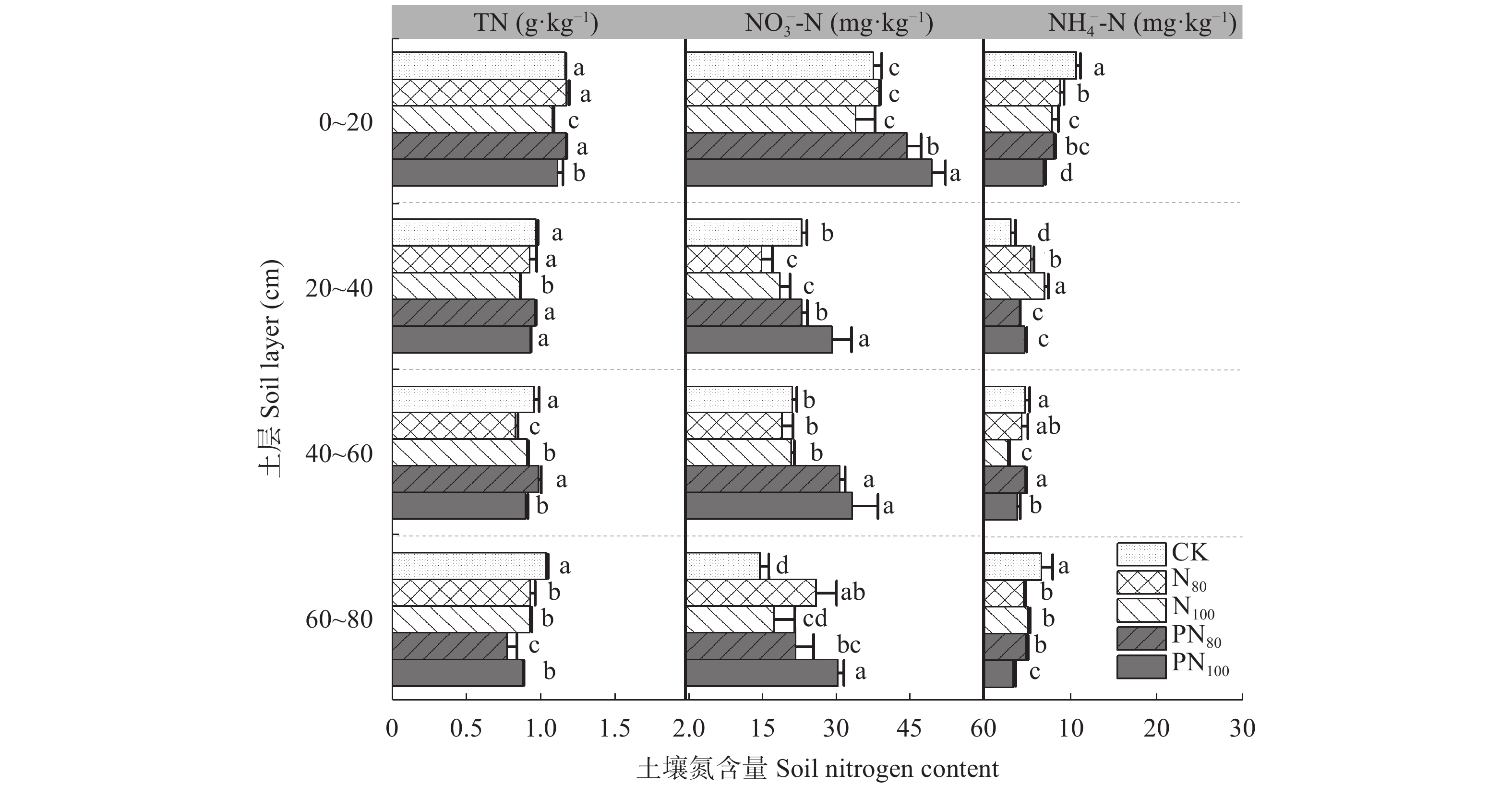
为探究减氮下调理剂对土壤养分及莜麦产量的调控作用, 以自主研发的新型液态调理剂为研究对象, 设置不施肥(CK)、80%氮肥(N80)、常规施氮(N100)、调理剂+80%氮肥(PN80)、调理剂+常规施氮(PN100) 5个处理, 分析不同氮肥用量添加调理剂对冀西北地区土壤物理性状、不同土层碳氮组分含量及莜麦产量的影响。结果表明, 与常规施氮(N100)处理相比, PN80处理的土壤含水量、田间持水量、孔隙度分别显著提高8.72%、8.22%、17.68% (P<0.05), 土壤容重显著降低9.06% (P<0.05)。对土壤有机碳、全氮及其组分研究表明, PN80处理较N100显著提高0~60 cm土层的有机碳、全氮、硝态氮和20~60 cm土层活性有机碳、微生物量碳, 分别提高4.97%~20.06%、8.43%~11.66%、23.10%~44.96%和11.95%~40.49%、11.43%~40.42% (P<0.05)。不同处理对莜麦养分及产量的影响差异较大, 其中PN80处理效果最为显著。与N100相比, PN80的莜麦全氮、全磷、全钾含量及作物产量分别显著提高12.93%、15.16%、3.69%、18.73% (P<0.05)。同时, 与N100相比, N80在减氮20%的情况下显著降低了莜麦NPK吸收量, 但并未造成莜麦减产。综上所述, 较常规施肥措施, 减氮(80%氮肥)添加调理剂可以改良土壤性状, 增加土壤碳组分含量, 减少硝态氮淋溶的风险, 进而提高莜麦植株的养分及产量, 这对实现农业绿色发展, 减少肥料投入对环境产生的影响, 提高氮肥利用率具有重要意义。
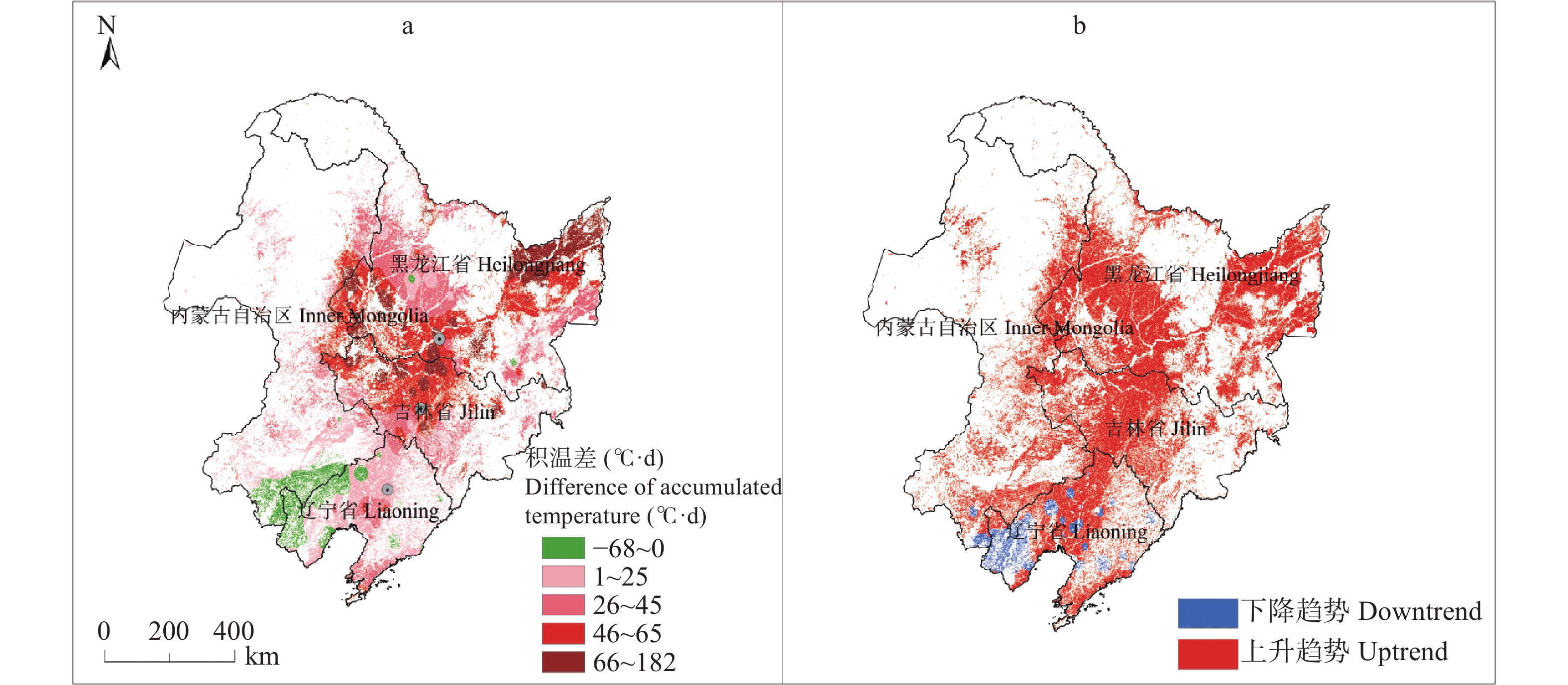
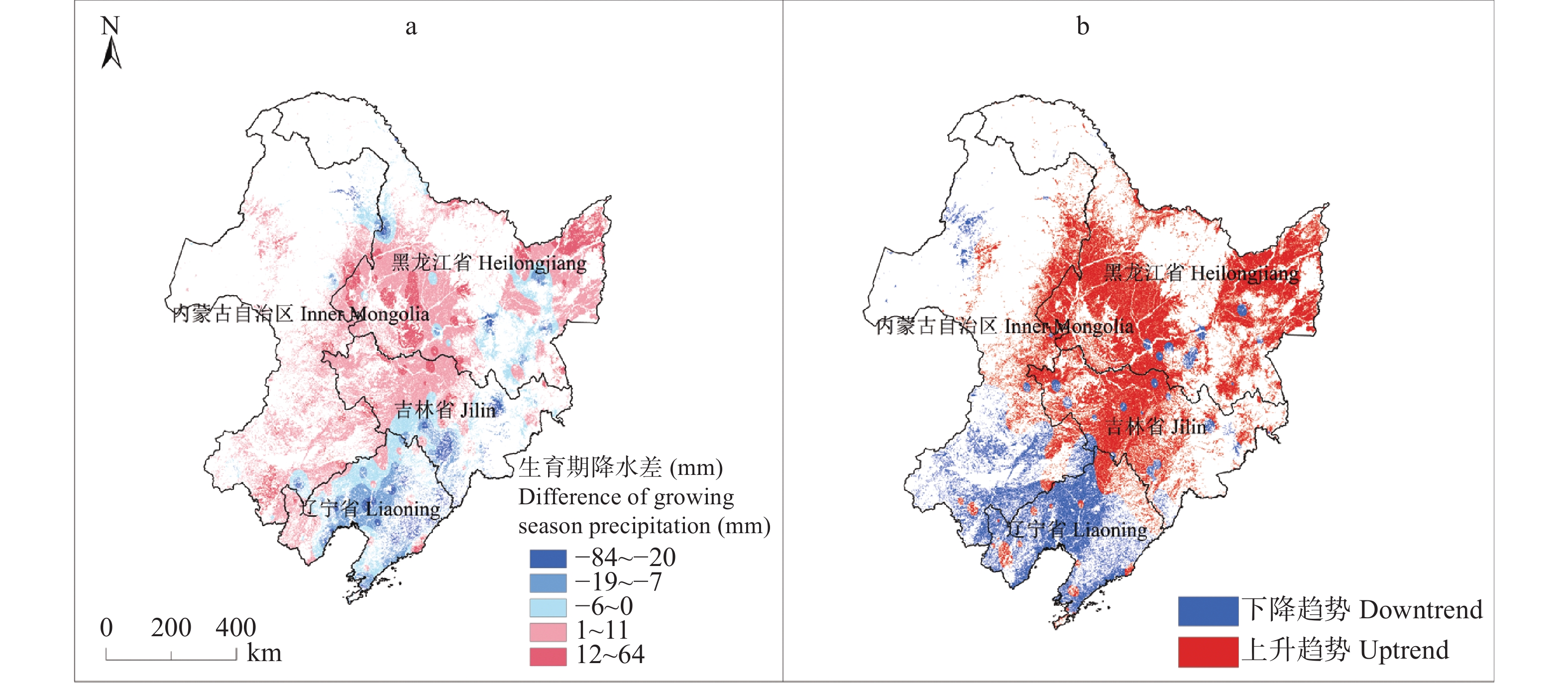
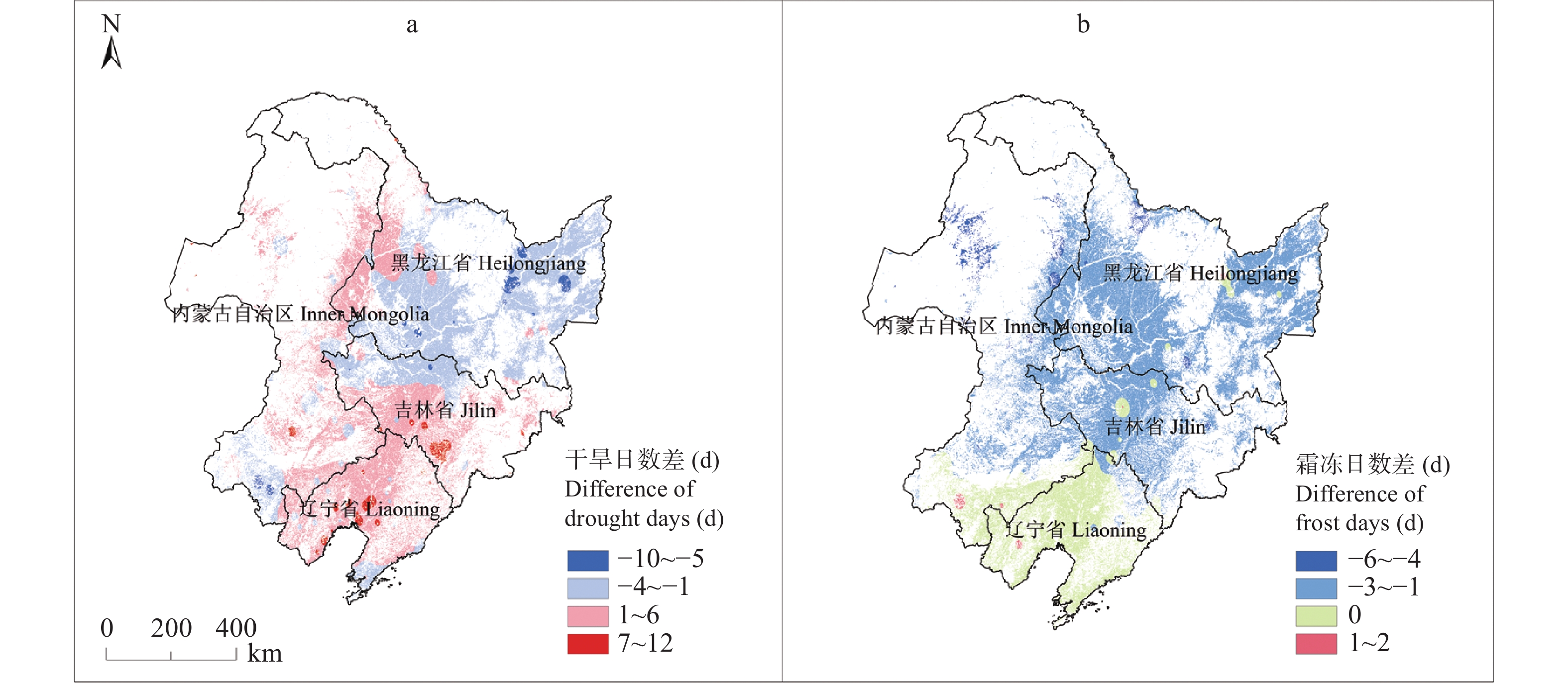
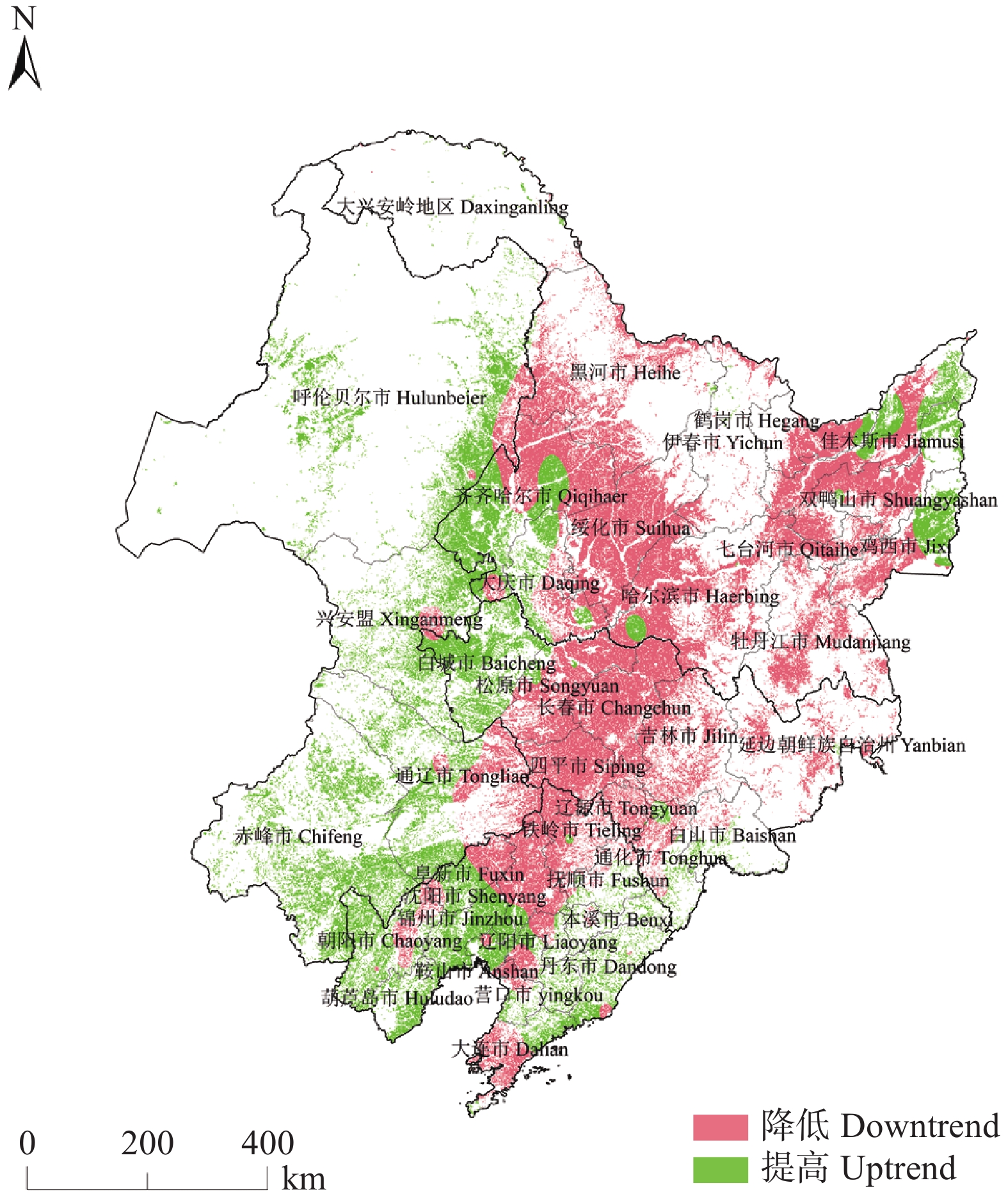
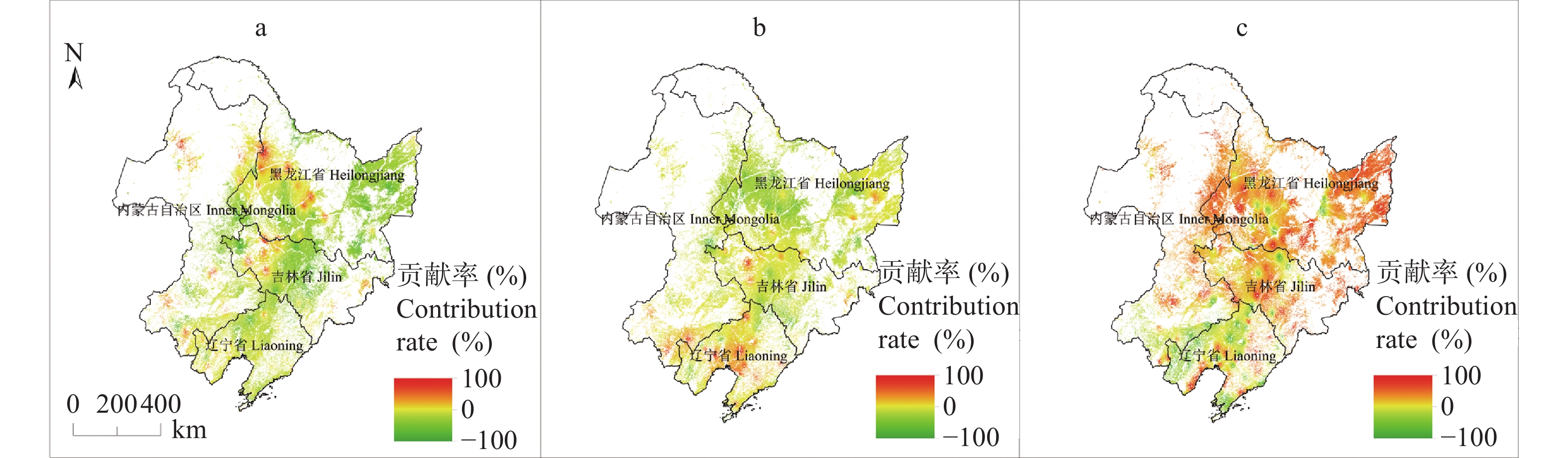
东北地区是我国大豆主产区, 该地区深受气候变化影响。厘清气候变化背景下的农业气候资源、农业气象灾害、大豆气候种植适宜性变化对指导大豆生产和扩种具有重要意义。通过利用最新气候态(1991−2020年)与旧气候态(1981−2010年)的农业气候资源、农业气象灾害, 分析了大豆种植气候适宜度的时空变化。结果发现: 1)内蒙古东部、黑龙江西南部和东北部、吉林中西部等地大豆生长季≥10 ℃活动积温增加26~65 ℃∙d, 局部地区增加66~182 ℃∙d; 1981−2020年整个时间段, 东北大豆的种植区生长季积温大部呈上升趋势。大豆生长季降水和日照变化呈现空间差异。2)东北地区大部大豆生长季霜冻害日数减少; 黑龙江大部和吉林西北部大豆生长季干旱天数减少, 吉林中东部和辽宁大部干旱天数增加。3)内蒙古东部、黑龙江西部、吉林西部、辽宁西部和东部大豆种植气候适宜度提高, 黑龙江中部和东部、吉林中部和东北部气候适宜度略下降。4)气象因子对大豆气候适宜度变化贡献率分析表明, 日照变化是大豆气候适宜度变化的主要因素, 其次是积温变化, 降水变化对大豆气候适宜度变化贡献率最小。最新气候态下东北地区大部大豆种植气候条件变好, 东北地区可以充分利用温度资源, 适当调整播期和品种, 但从积温上升幅度看, 不宜采取过激的跨积温带盲目种植。

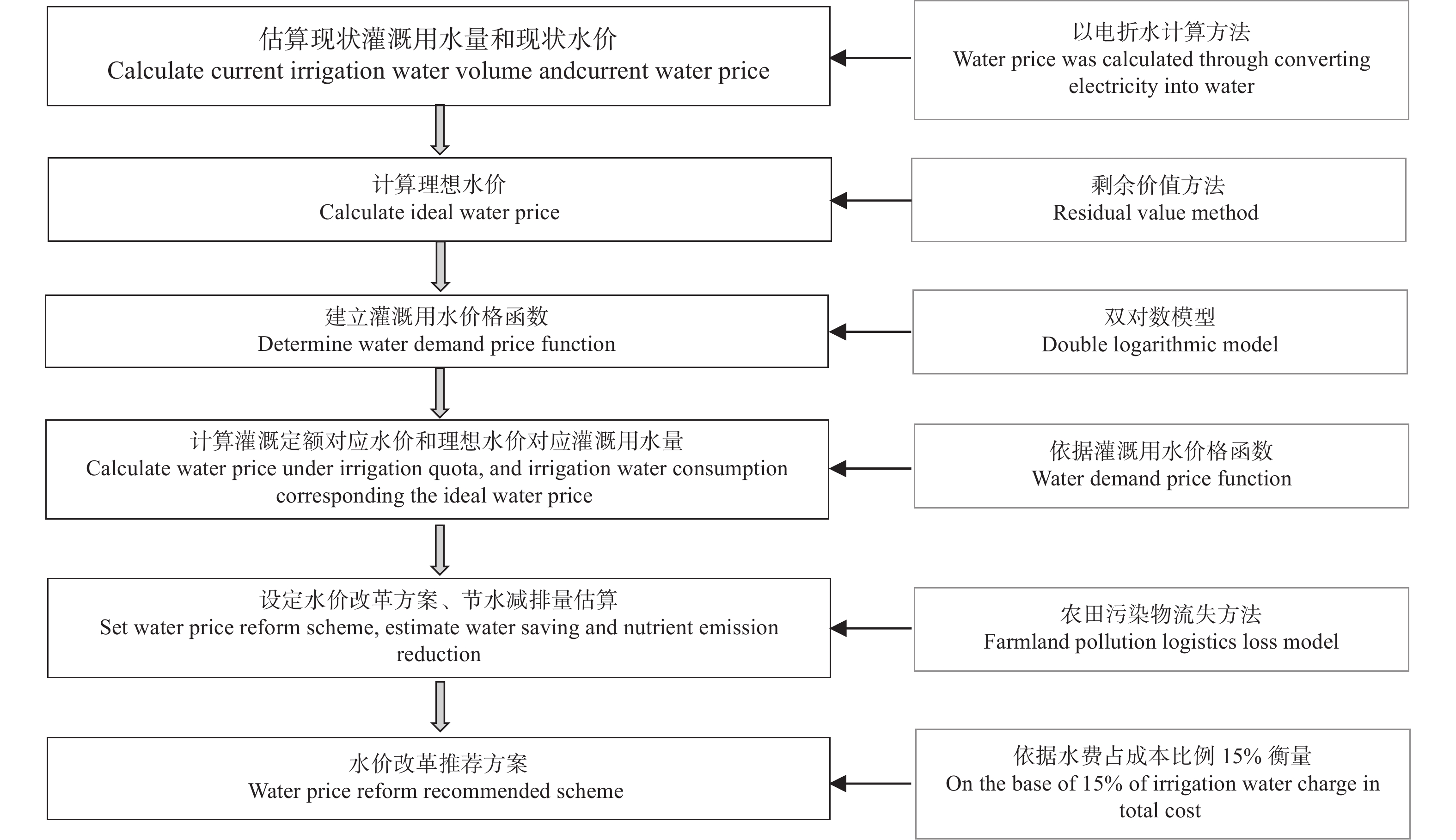
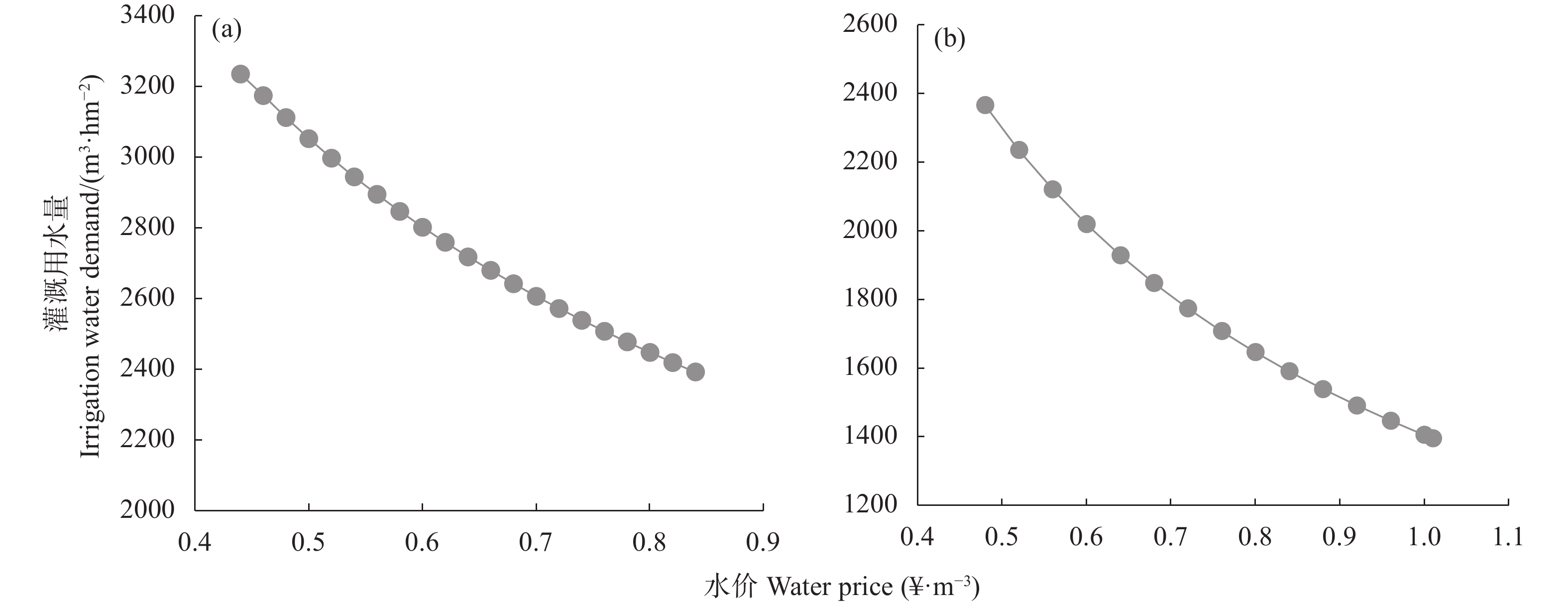
研究并提出合理的水价调整方案是农业水价综合改革的需求。本文提出地下水灌区水价确定及其节水减排估算方法, 采用“以电折水”方法计算现状灌溉用水量与现状水价, 运用双对数模型建立灌溉用水价格弹性函数, 依据剩余价值方法计算理想水价, 借鉴农田营养物流失模型计算节水的营养物减排量。以河北省南皮县为例进行实证分析, 研究表明: 1)根据问卷调研数据与“以电折水”计算方法, 获得小麦和玉米现状水价分别为0.44 ¥∙m−3 和0.48 ¥∙m−3。2)根据现状水价、现状灌溉用水量与双对数模型,获得小麦和玉米水价弹性系数分别为−0.47和−0.71。3)灌溉定额对应的理论水价小麦和玉米分别为0.52 ¥∙m−3和0.77 ¥∙m−3, 理想水价分别为0.84 ¥∙m−3和1.01 ¥∙m−3。4)以理论水价作为推荐方案, 该方案下水费占成本比例低于15%, 小麦和玉米水价提升空间分别为0.08 ¥∙m−3和0.29 ¥∙m−3, 节水潜力为235.05 m3∙hm−2和682.80 m3∙hm−2; 氨氮、总氮和总磷的减排量小麦为5.2~19.2 g∙hm−2、52.7~195.4 g∙hm−2和4.6~16.9 g∙hm−2, 玉米为18.5~27.6 g∙hm−2、189.1~281.2 g∙hm−2和16.3~24.3 g∙hm−2。农业水价综合改革是一项系统工程, 需要相关配套政策的支持, 建议在节水技术采纳、土地流转和规模化经营、灌溉定额管理制度等方面, 给予政策倾斜和相关的激励政策, 以利于农业水价综合改革的推进。
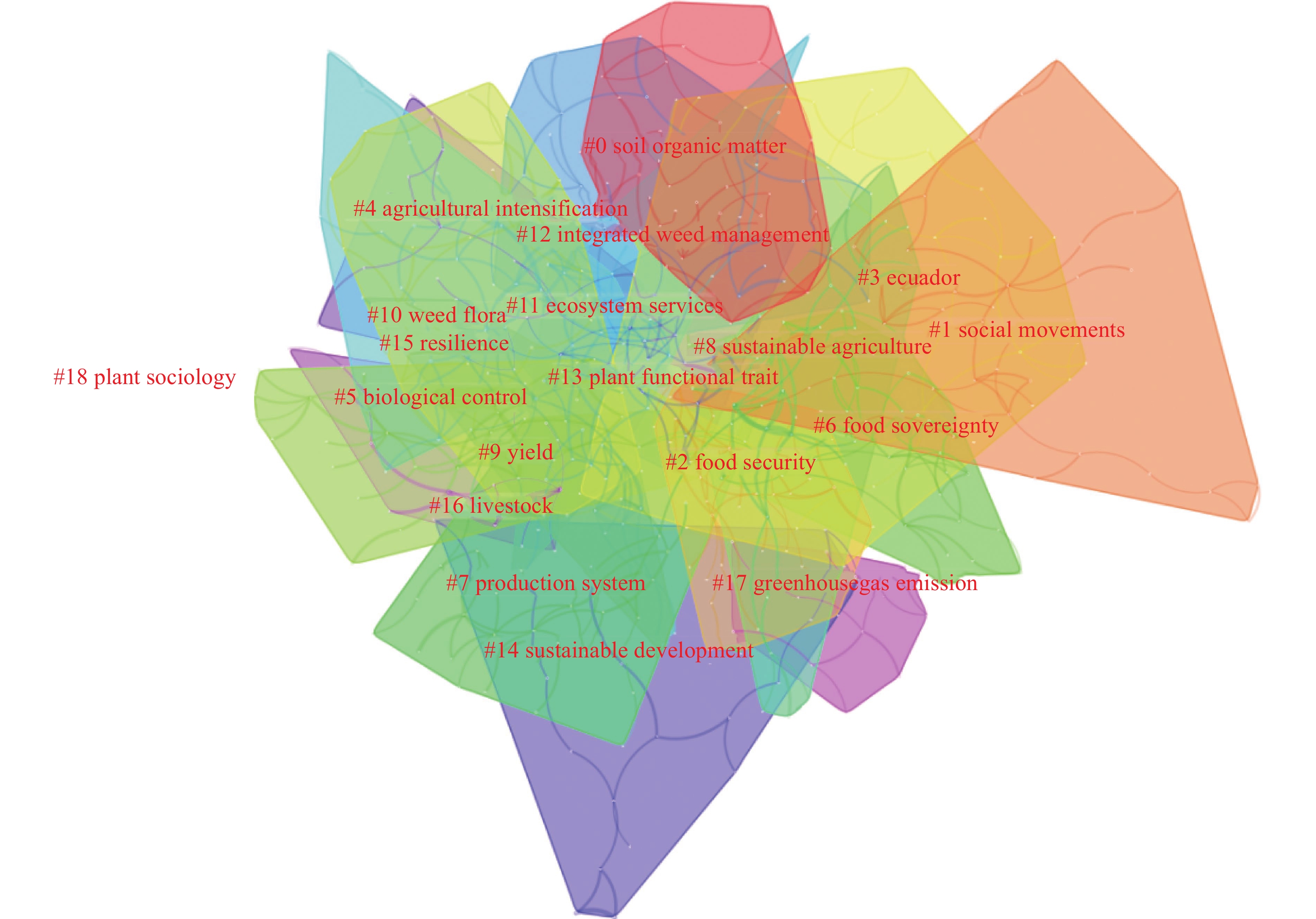
农业生态学作为生态学和农业科学的交叉学科, 有助于优化传统农业体系和生态农业实践。为厘清国际农业生态学的发展现状和研究趋势, 本文借助CiteSpace对2012—2021年Web of Science核心数据库的国际农业生态学论文进行计量分析, 研究发现: 第一, 国际农业生态学的热点议题主要关注作为农业生态系统的整体表现及其评价的研究、以生态农业实践为基础的科学研究以及关于生态农业社会运动与社会影响的研究等方面。第二, 国际农业生态研究人员在农业生态学领域有很多基础性和创新性研究, 主要围绕农业生态转型与生态农业产业形态、生态农业社会运动以及如何调节农业生态系统内部诸生态因子建构可持续农业生态体系展开。第三, 国际农业生态学研究的前沿趋势不仅聚焦于涉及自然学科的生物多样性保护、景观生态安全格局、农业种植体系与农药污染等内容, 还涉及社会管理相关领域的知识农民培育、农业教育和参与式治理模式等方面研究。未来国内农业生态学的发展可在立足国情的基础上, 加强国际交流合作, 同时, 紧随国家发展战略, 从而推动新型农业技术的创新发展。

实现农业绿色生产有赖于小农户生产方式的转变。相比以往研究多聚焦于人工施肥, 本研究关注农户使用机械后, 通过机械搭配使用化肥对化肥施用量以及施肥均匀度的影响。利用2017年中国农村家庭追踪调查(CRHPS)数据, 通过OLS以及处理效应模型探讨农业机械化对小麦化肥使用影响, 并关注到不同机械使用来源化肥施用差异。研究结果表明: 整体而言, 使用机械并不会降低小麦化肥使用量, 但能够缩小农户间施肥差异。分环节来看, 只有机播环节能够降低化肥施用量以及施肥不均等, 这主要是因为小麦在机播环节施肥占比较高。区分农户农业机械使用来源差异后, 发现相比农户自购机械, 购买机械服务每公顷能够减少150.45 kg化肥施用量并提升施肥均匀度。其中, 购买机播服务同自购机械播种相比, 每公顷能降低213.45 kg施肥量并提升0.32个单位施肥均匀度。此外, 农户采用机播可使每公顷化肥投入下降283.95元, 购买机械服务则可使每公顷化肥投入平均下降394.50元, 购买机播服务能降低352.80元。这说明, 农业机械化能够实现化肥减量并降低农户间施肥不均等, 且主要通过服务外包实现。本文结论有助于扩展理解农业机械化与现代农业之间的关系, 从而为我国农业绿色转型以及可持续发展提供有力支撑。
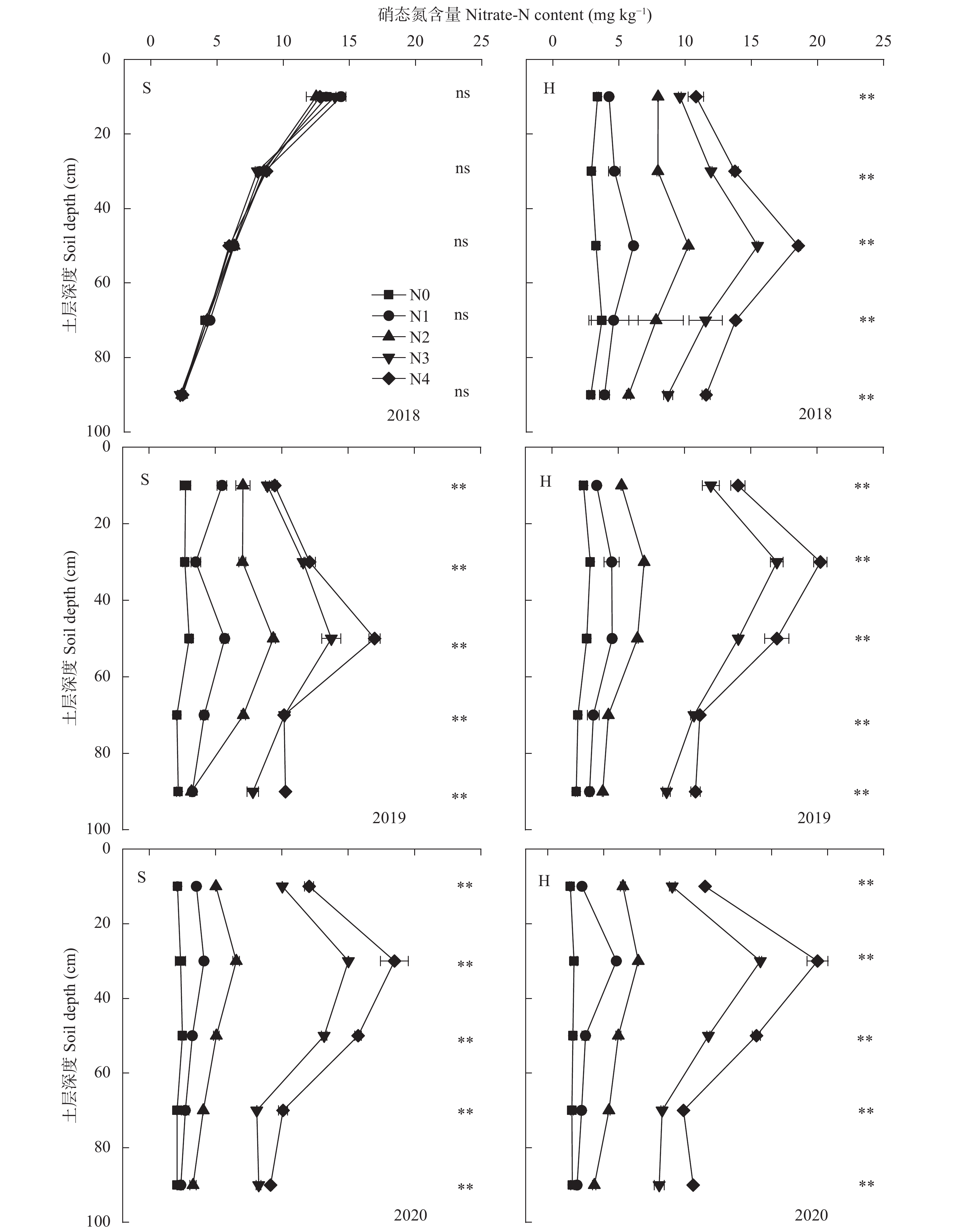
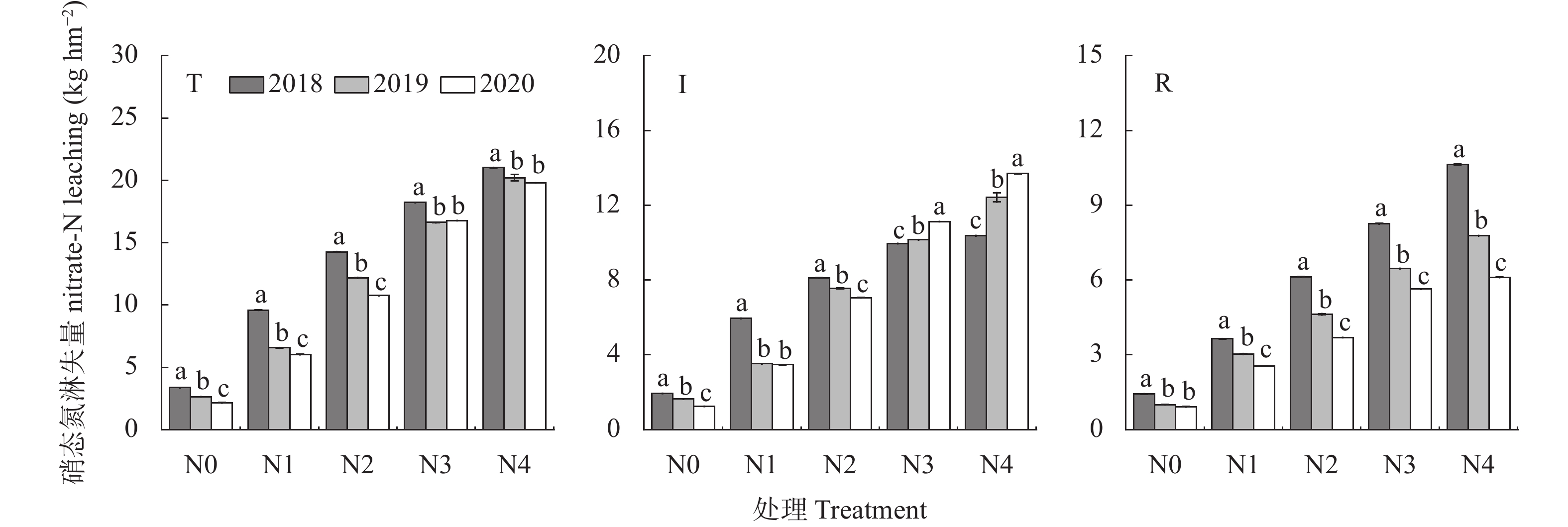
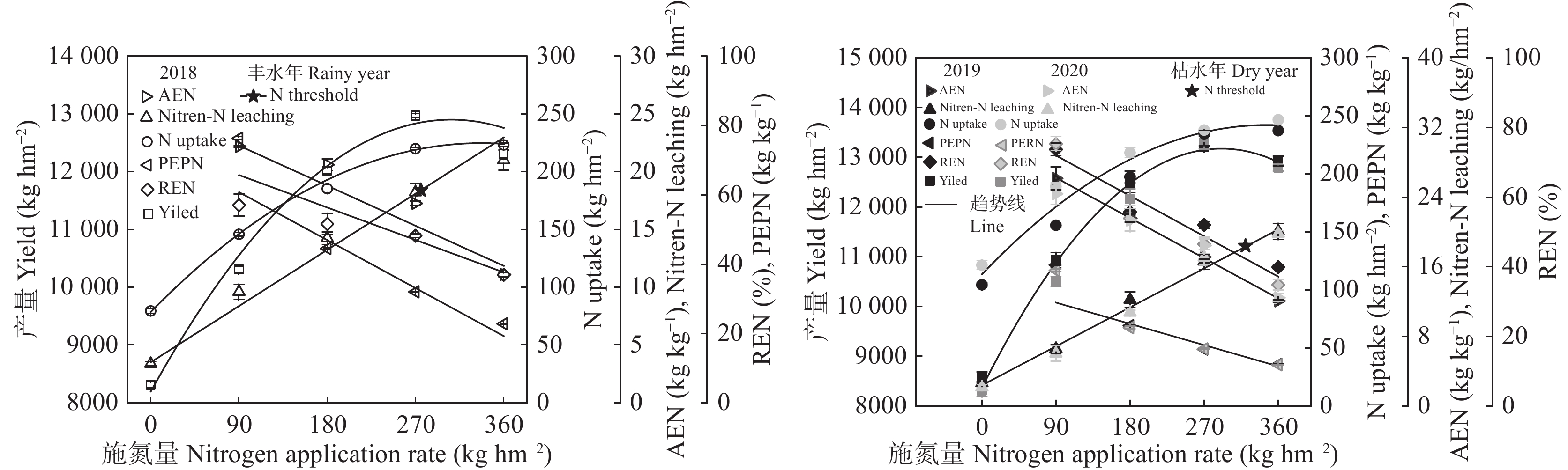
为寻找满足宁夏地区滴灌条件下不同降水年型间科学施肥模式, 缓解不合理施氮导致的资源浪费、黄河水质下降和地下水污染等问题, 本研究于2018—2020年在宁夏平吉堡农场开展氮梯度试验, 分析不同降水年型下不同施氮处理土壤硝态氮残留和淋溶量以及对滴灌玉米氮素吸收利用和产量的影响。结果表明: 土壤硝态氮含量峰值与降水量密切相关, 丰水年(2018年)硝态氮残留量峰值在40~60 cm土层, 枯水年(2019年和2020年)在20~40 cm土层; 不同降水年型份间土壤硝态氮残留量和淋失量均随施氮量的增加而增加, 且降水量显著影响硝态氮淋失量; 丰水年由降水因素导致的硝态氮淋失量占总淋失量的50.62%, 枯水年占总淋失量的34.82%。回归分析结果表明, 不同降水年型间玉米产量随施氮量呈先上升后下降的趋势, 均在N3处理 (施N量为270 kg∙hm−2)下达最高产量, 且N3处理的产量和吸氮量与N4处理(施N量为360 kg∙hm−2)无显著差异; 丰水年N3的氮肥利用率、氮肥农学利用率和氮肥偏生产力比N4分别提升11.38%、6.16 kg∙kg−1和13.85 kg∙kg−1, 枯水年分别提升12.10%、5.06 kg∙kg−1和15.00 kg∙kg−1。综合考量不同降水年型间0~100 cm土层硝态氮分布特征和硝态氮淋失量及施氮处理下的产量、氮素吸收利用, 推荐宁夏引黄灌区滴灌玉米不同降水年型下施氮量在270 kg∙hm−2时较适宜, 丰水年施氮最大阈值为275.59 kg∙hm−2 , 枯水年施氮最大阈值为320.20 kg∙hm−2。
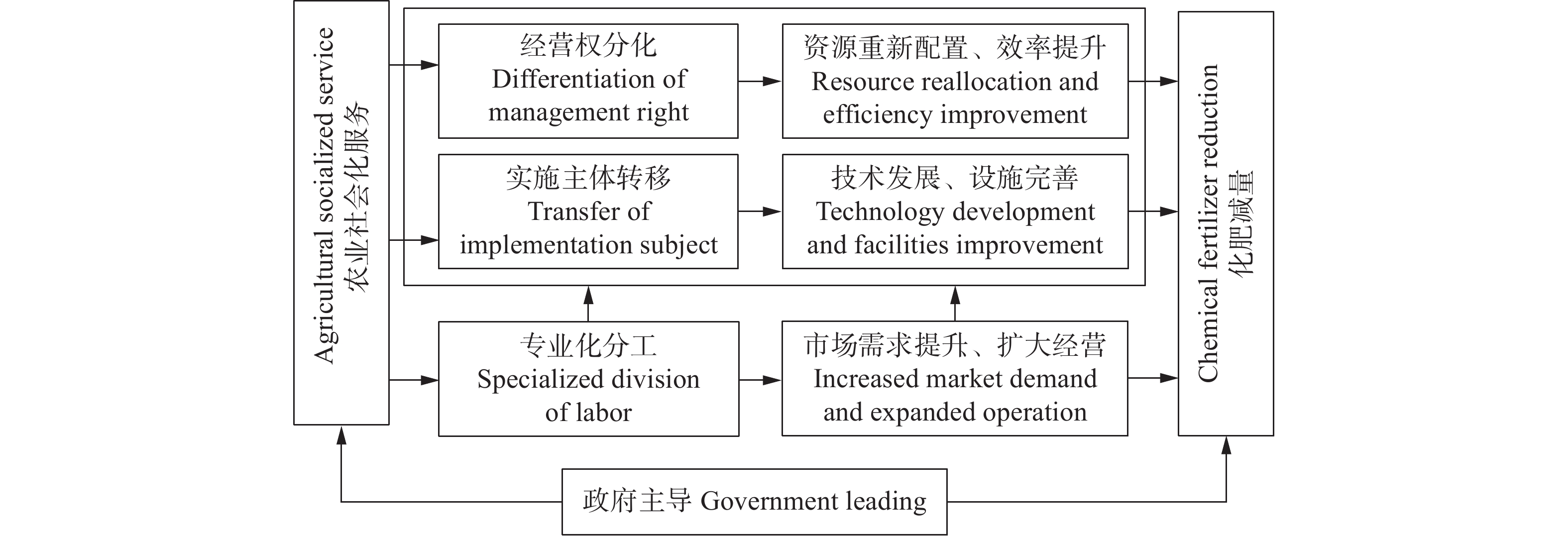
实现化肥减量是我国农业走向高质量发展的重要举措, 也是实现绿色化农业发展的关键所在。基于专业分工理论, 利用2010—2020年统计年鉴数据, 采用中介调节模型和工具变量模型实证分析了农业社会化服务对化肥减量的作用机制。研究发现, 农业社会化服务对化肥使用总量和单位面积使用量具有显著负向影响, 即即每增加1亿元的农业社会化服务产值能分别降低0.055万t的化肥使用总量和0.443万t的单位面积使用量。中介和调节分析发现, 农业社会化服务通过资源配置和科技进步两条路径实现化肥减量, 分工深化和政府主导对该路径分别对该路径存在0.039的促进作用和−0.017的抑制作用。进一步研究发现, 农业社会化服务可以显著降低服务规模小、农业发展程度高和东部地区省份的化肥使用量, 而对中西部地区省份化肥减量未能发挥作用。其中, 监管体系的缺失和分工深化的不足是目前我国农业社会化服务面临的主要问题, 也是阻碍化肥减量实现的重要因素。
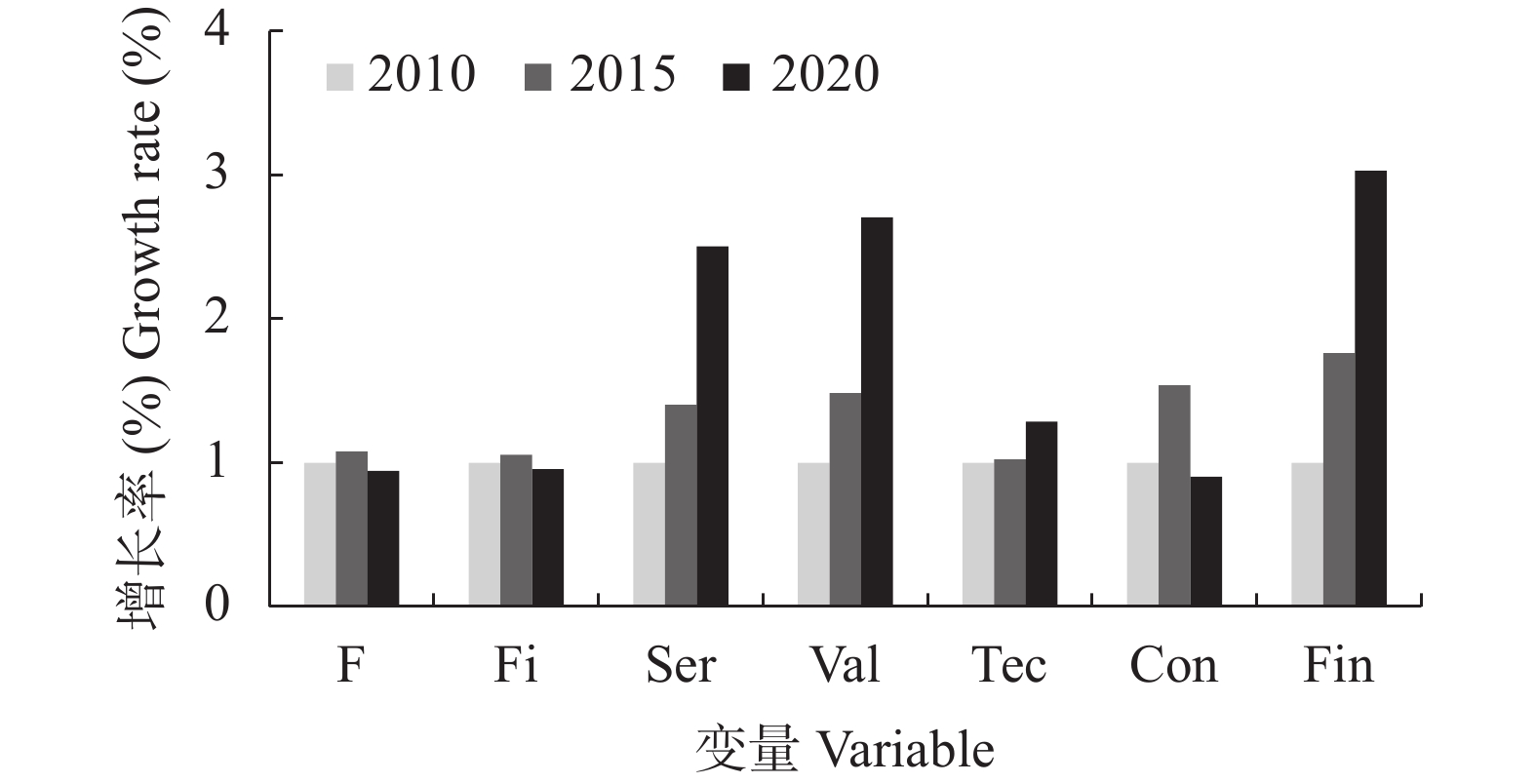
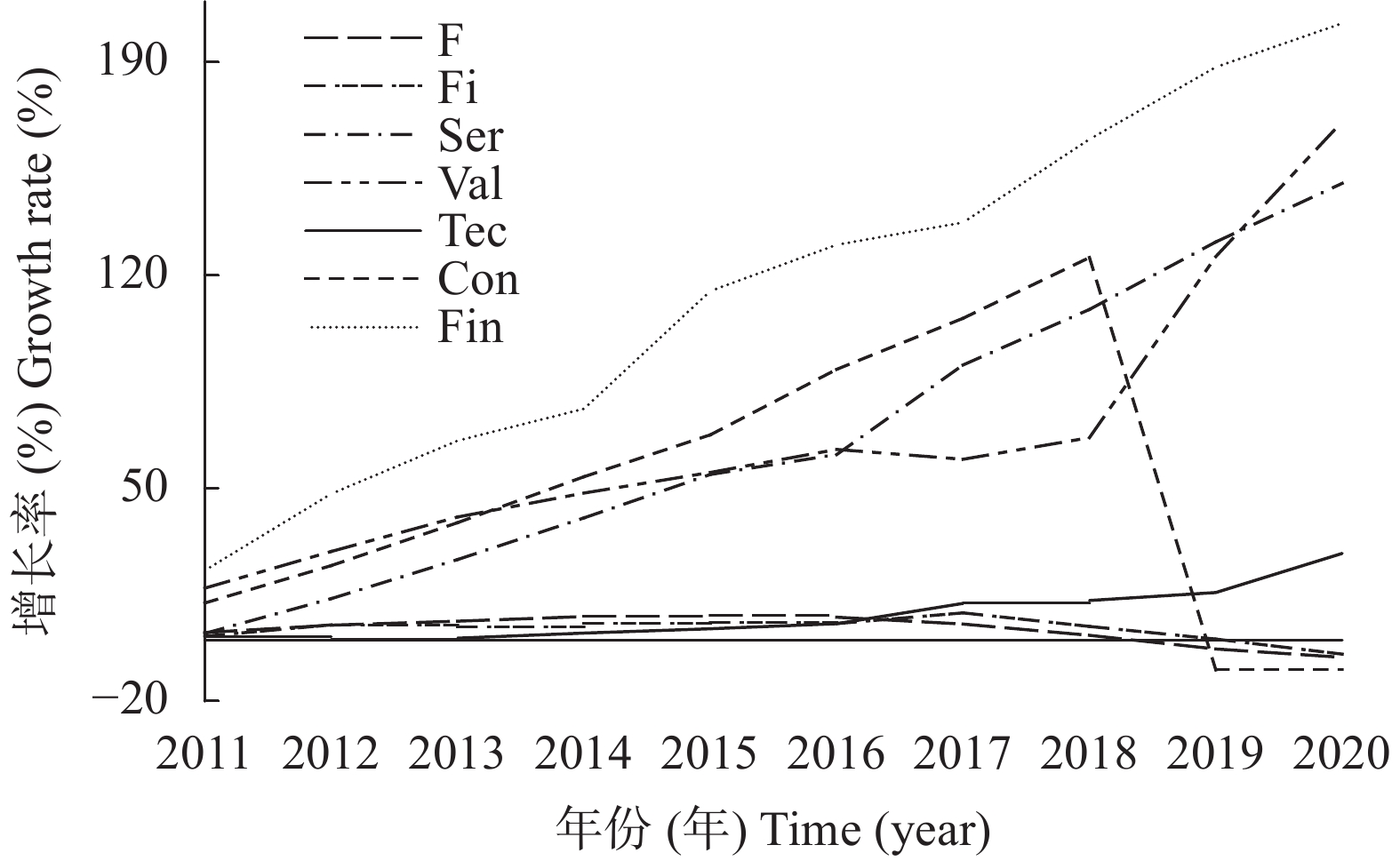
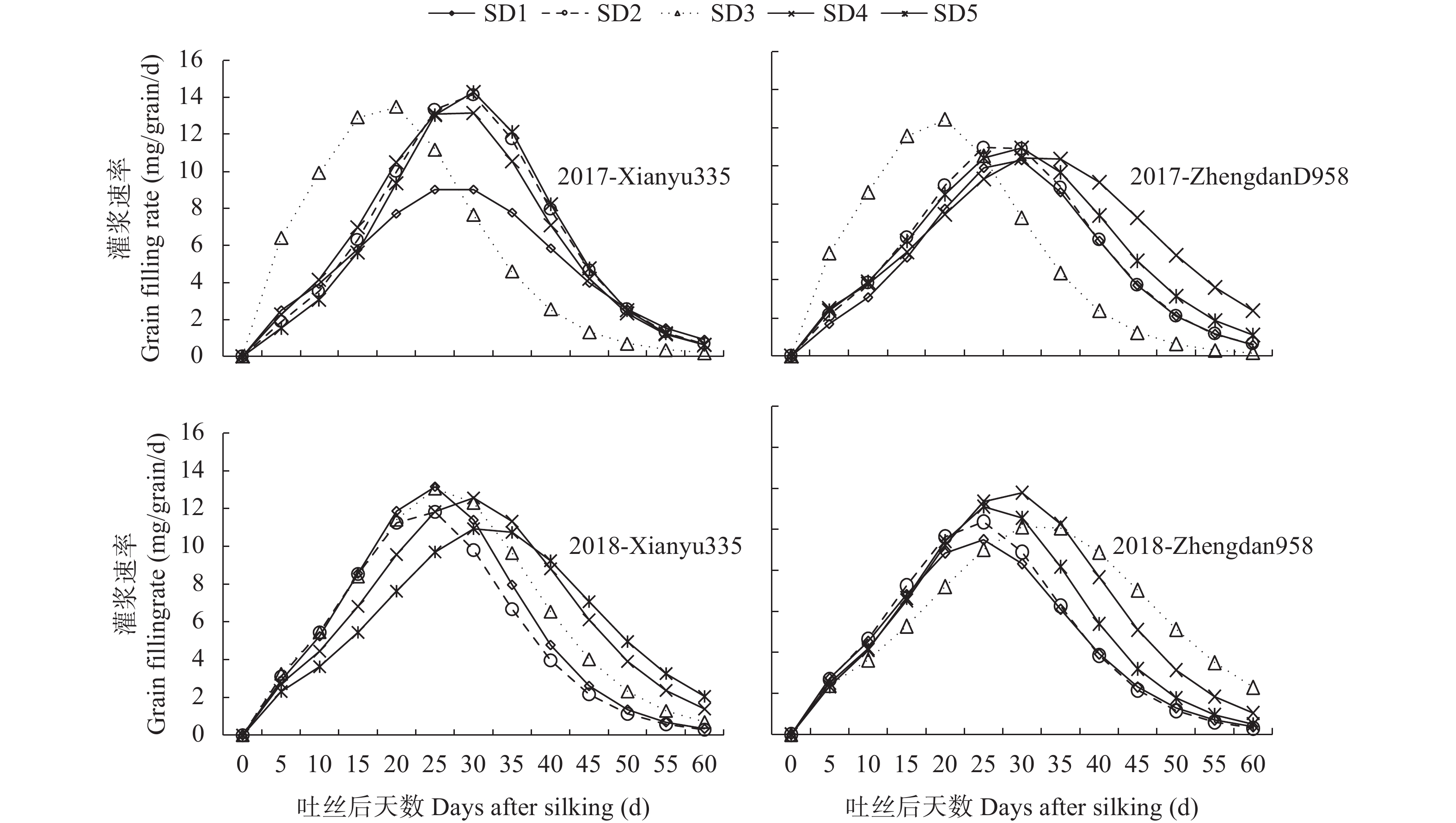
本文通过研究在限水灌溉条件下, 不同播期对一作玉米产量、产量性状和籽粒灌浆特性的影响及阶段气象因子与产量、产量性状及灌浆参数的相关关系, 探讨了播期对一年一作玉米产量的影响机制, 为低平原区高产高效生产提供了数据支撑。试验采用‘先玉335’和‘郑单958’ 2个玉米品种, 设置5个播期: 5月5日(SD1)、5月20日(SD2)、6月5日(SD3)、6月20日(SD4)和6月30日(SD5)。结果表明: 1)播期对产量影响显著, 且受品种和年型综合影响。随播期的延后, 产量呈现出先增加后降低的趋势, 2年平均以SD1产量最低, SD4产量最高, SD3和SD4间产量差异不显著。SD4产量高的主要原因是穗粒数和百粒重较高。2)通径分析表明, 产量要素对产量的贡献相互影响, 其中对产量的直接作用最大的是百粒重。3)采用Logistic模型研究粒重变化特征(R2>0.98, P<0.01) , 粒重的大小由平均灌浆速率(V)和灌浆持续期(D)的乘积决定, 其中D对粒重的直接贡献最大。4)从气象因子对产量和百粒重的影响来看, 播种到吐丝的日平均气温(TAvsf)越高, 吐丝到成熟的日均温差TRAvfm越大, 产量越高; 吐丝后10 d≥35 ℃的天数(D1fa)越多, 吐丝到成熟的日均气温(TAvfm)越高,产量和百粒重越低; 其中TAvfm对产量和百粒重的直接贡献最大, 且各气象因子对产量和百粒重的作用相互影响。SD1粒重较低的主要原因是TAvfm高、TRAvfm小, 且D1fa的天数多, D较短。SD3和SD4 粒重较高的主要原因是TRAvsf较大, V、D较高。SD5虽然TRAvfm较大, 但因TAvfm较低, 总体D缩短, 最终粒重降低。5)从品种来看, ‘先玉 335’较‘郑单 958’产量高的主要原因是穗粒数和百粒重高, 且‘先玉 335’的V和V×D分别较‘郑单958’高0.19 mg∙grain−1∙d−1、0.73 mg∙grain−1∙d−1。这说明选用产量潜力大、灌浆速率高的品种, 在6月上旬到6月中下旬播种可优化生育期气象要素, 进而提高粒重和产量。
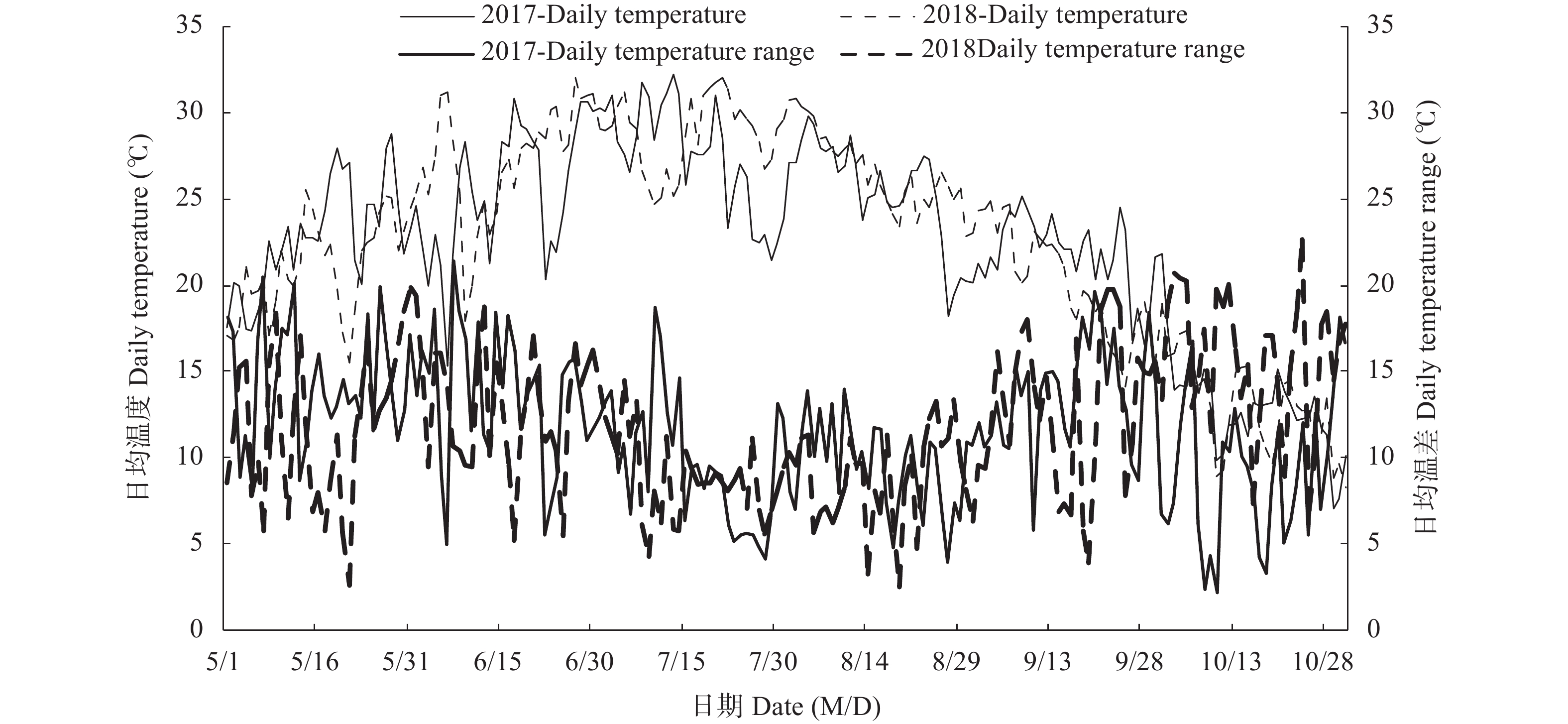
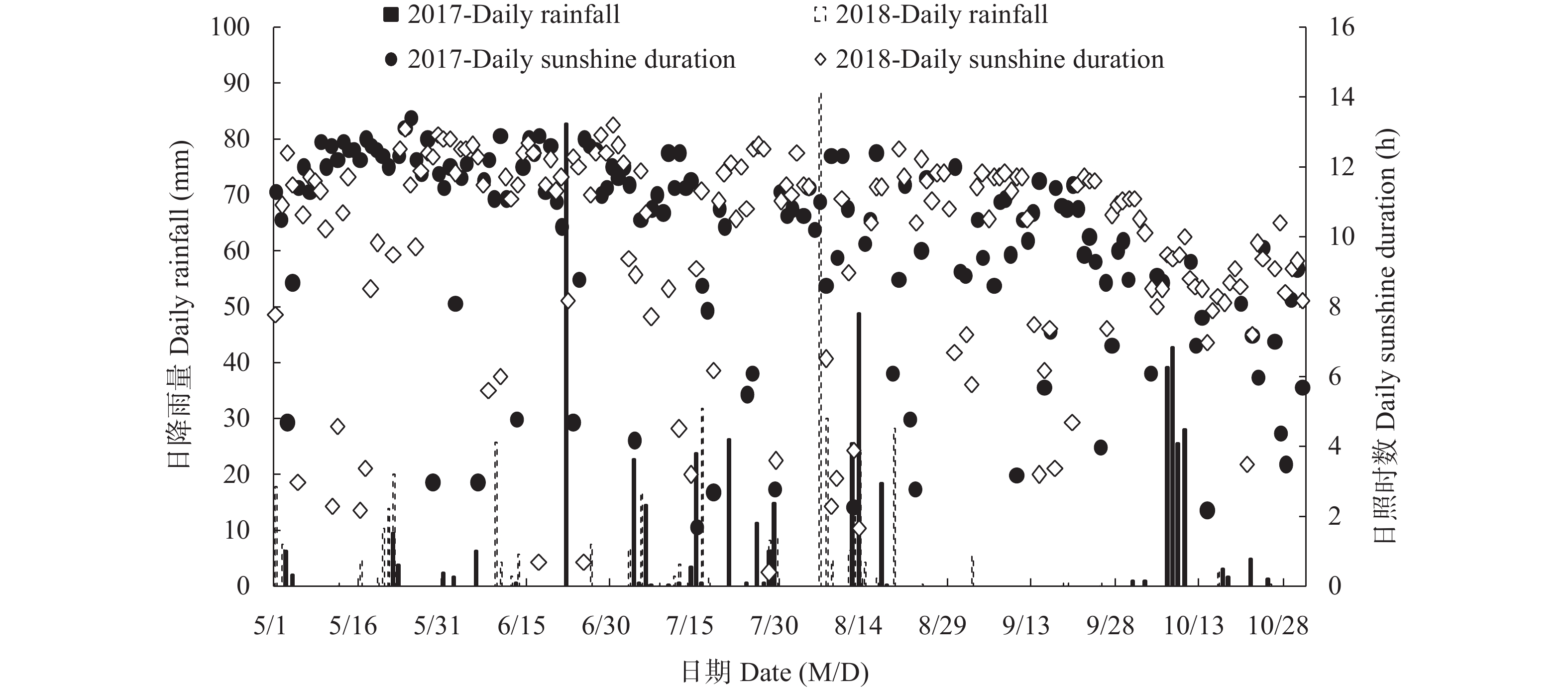
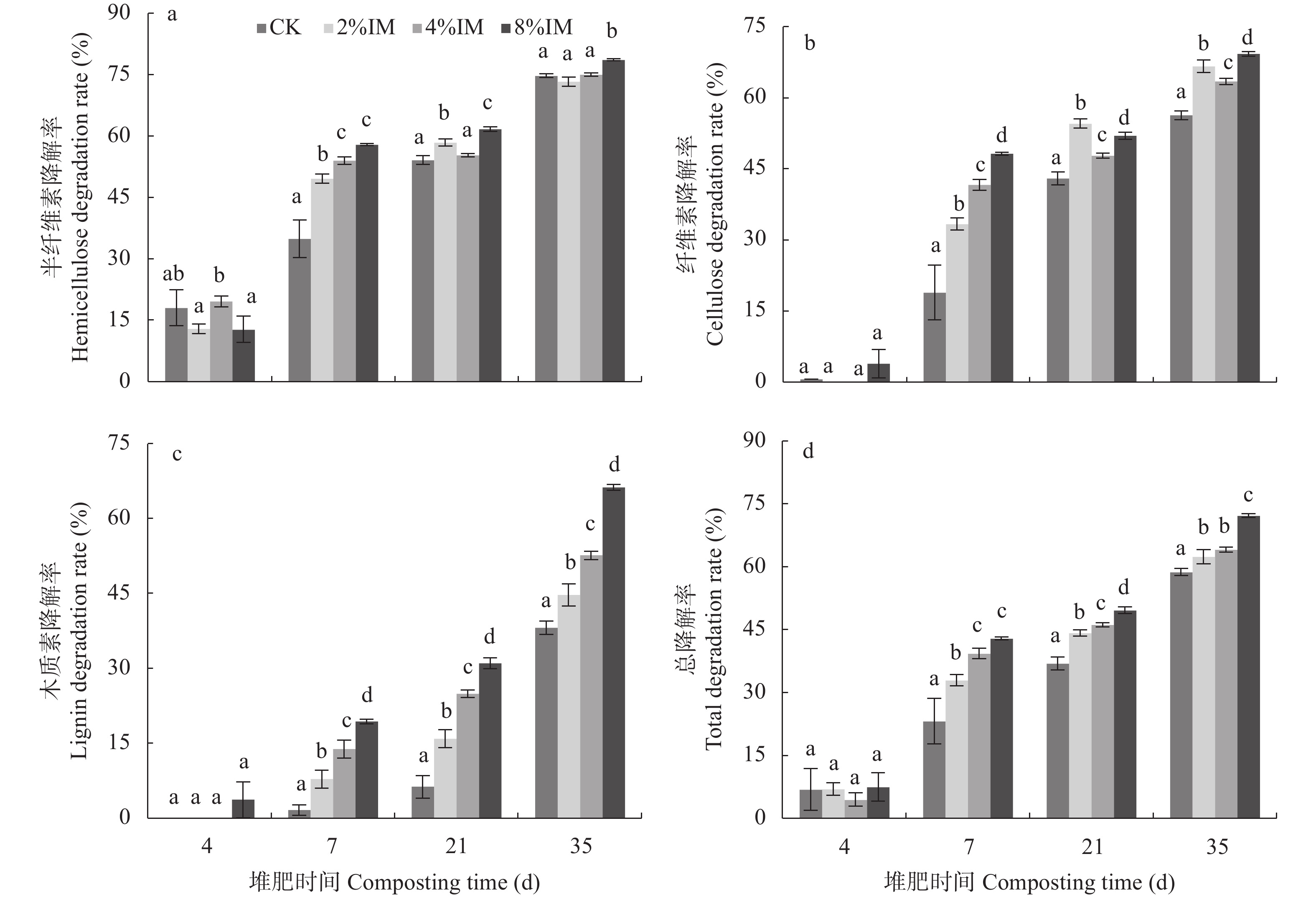
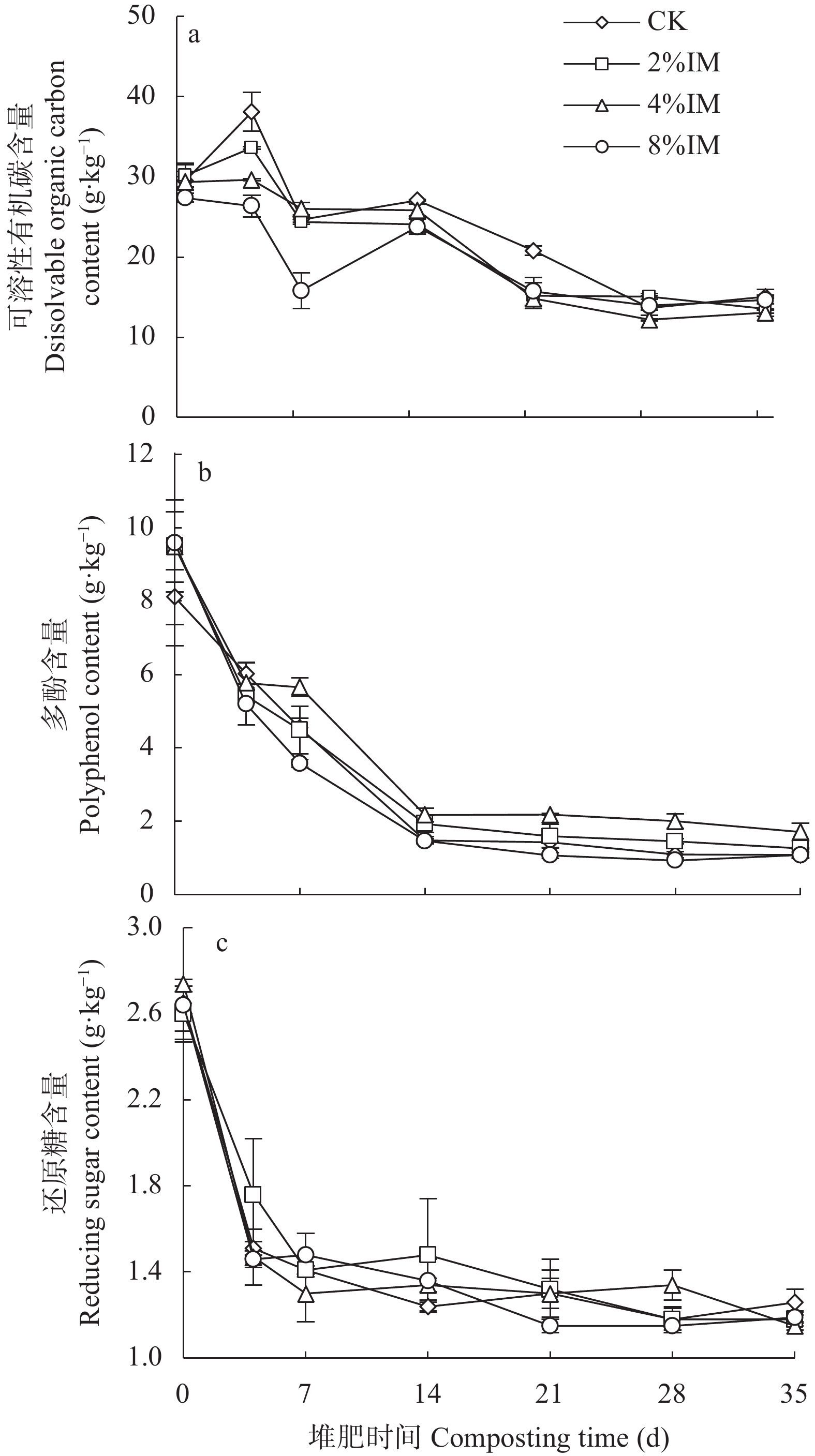
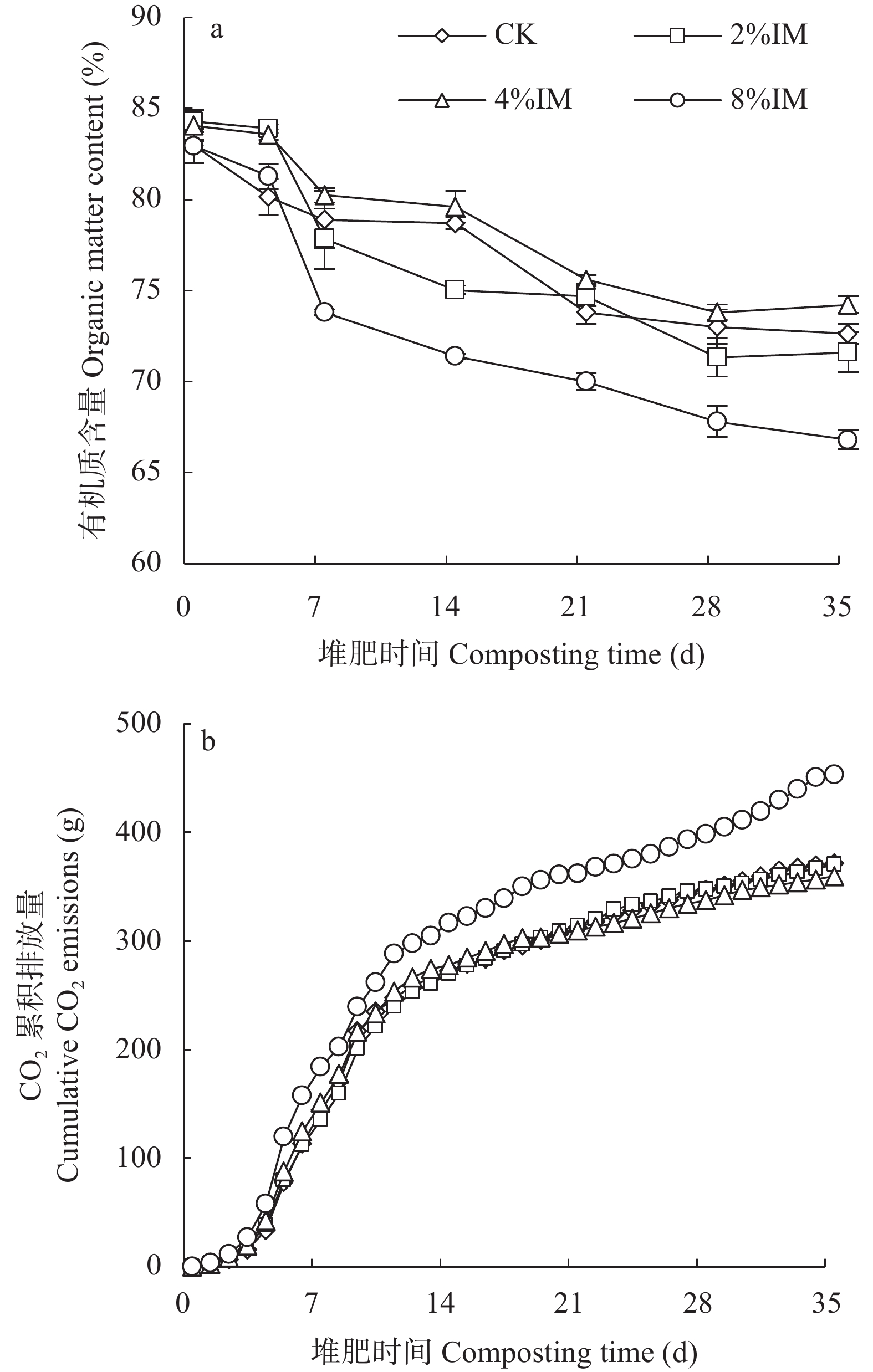
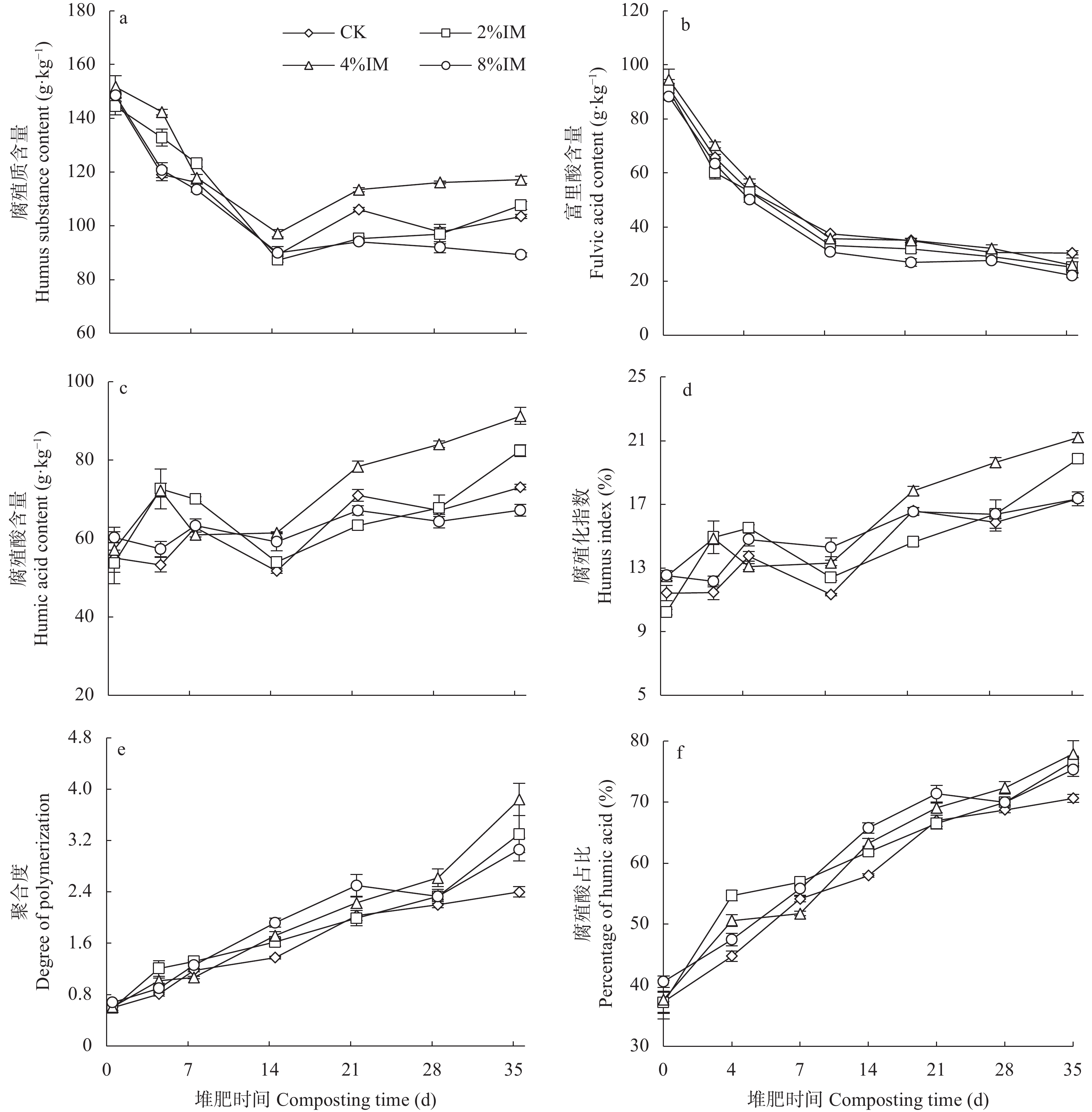
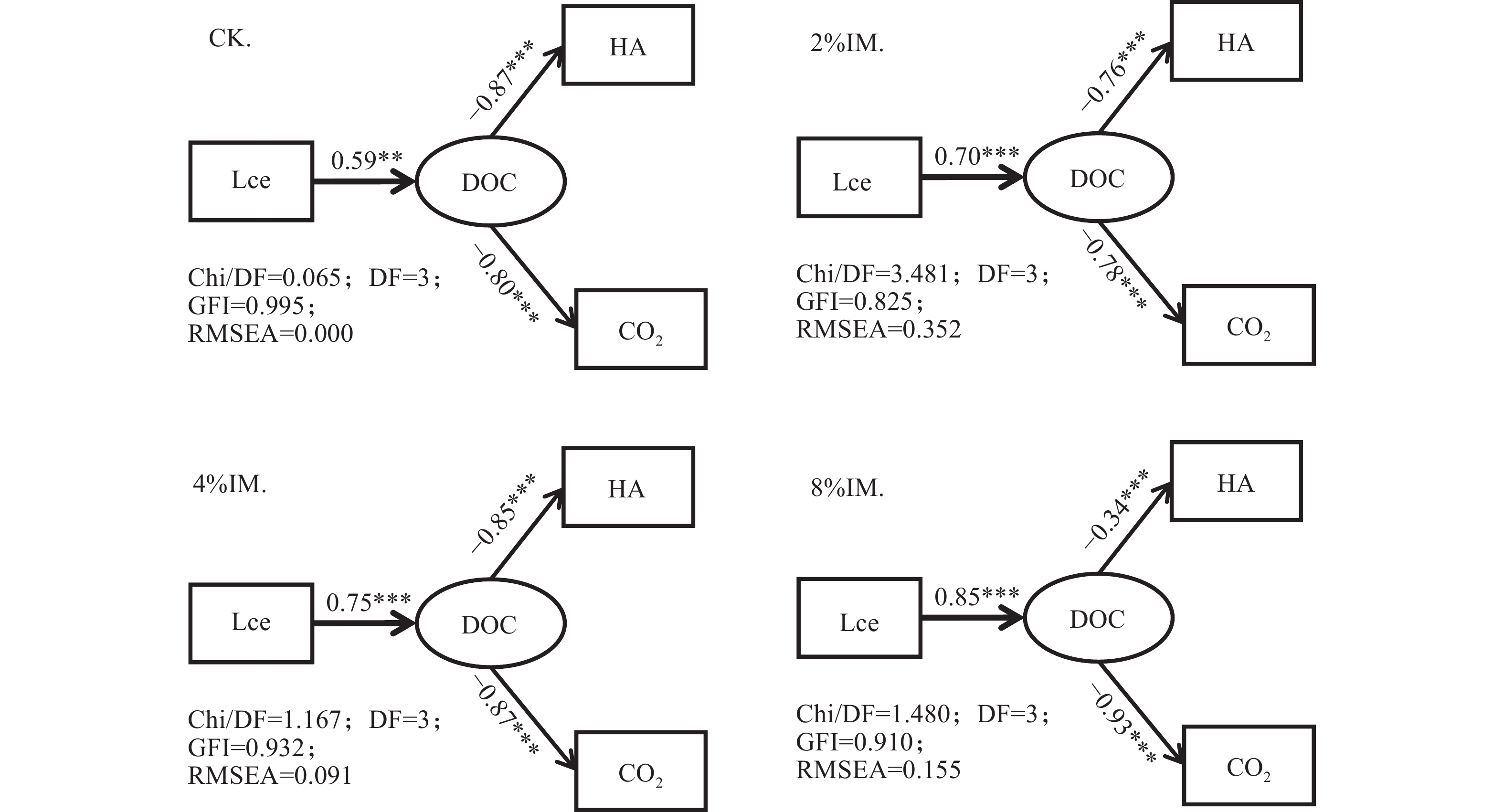
高木质纤维素含量制约了园林废弃物的堆肥化应用, 添加外源菌剂是加快木质纤维素降解的有效手段。为降低菌剂生产成本并提高接种效率, 本研究利用食品残渣(苹果渣、豆渣)代替常规碳氮源(葡萄糖、蛋白胨)进行木质纤维素降解菌的扩繁, 扩繁产品活菌数高达3.7×1010 cfu∙mL−1, 相较工业培养基增加了46.2%。探讨了不同降解菌接种量(0、2%、4%、8%, 干重计)对园林废弃物堆肥过程碳素转化的影响。结果表明, 接种处理显著增加了木质纤维素的降解(P<0.05), 2%、4%、8%接种处理(2%、4%、8%IM)的总木质纤维素降解率较CK分别提高6.3%、9.2%、23.0%, 其中8%IM处理加速了碳素的完全矿化, 导致腐殖质前体物(多酚、还原糖)被完全降解生成了CO2, 抑制了腐殖化的进行, 而4%IM处理在加快木质纤维素降解的同时促进了腐殖酸(HA)的合成, 其最终HA含量达到91.3g/kg, 较CK、2%IM、8%IM处理分别提高了24.9、10.7、35.8%。因此, 以食品残渣为培养基质可完全实现木质纤维素降解菌的生长扩繁, 同时, 4%接种量更有利于园林废弃物堆肥腐殖化的进行和碳素的保存, 本研究为多源废弃物高效协同处理提供理论依据。
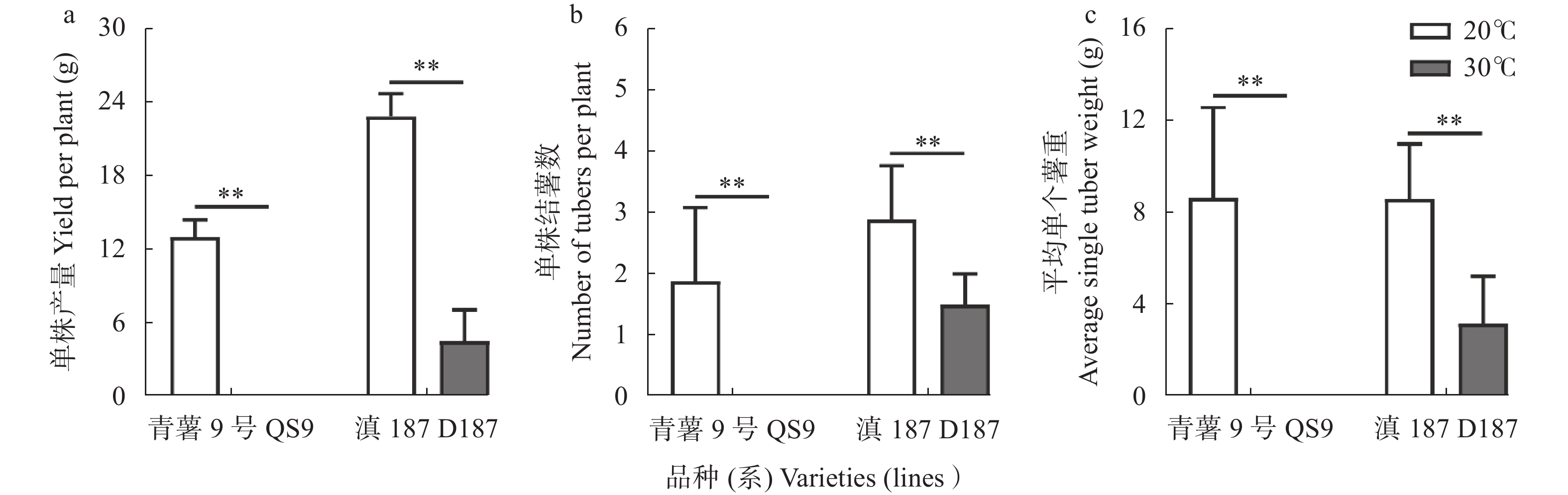
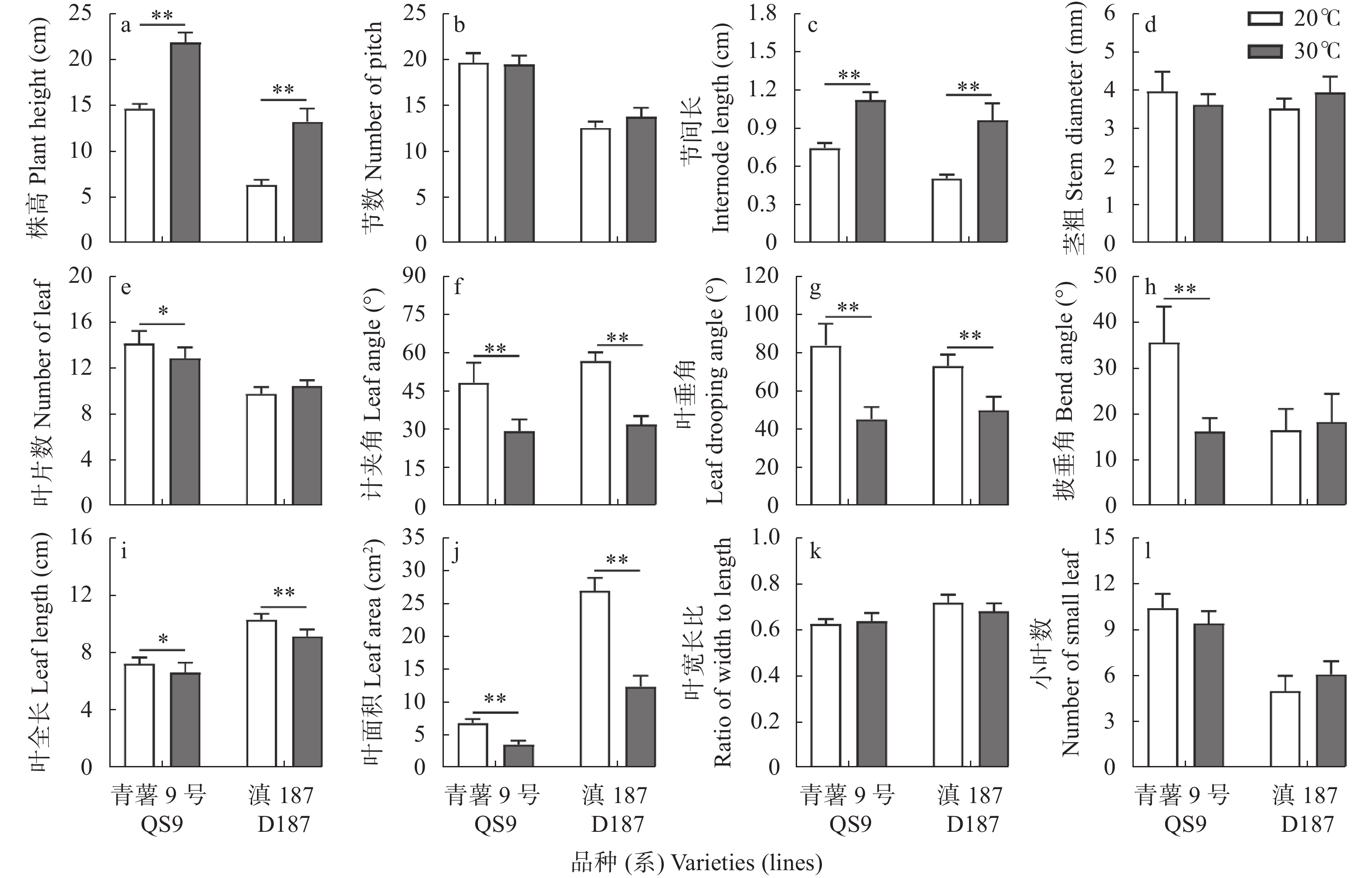
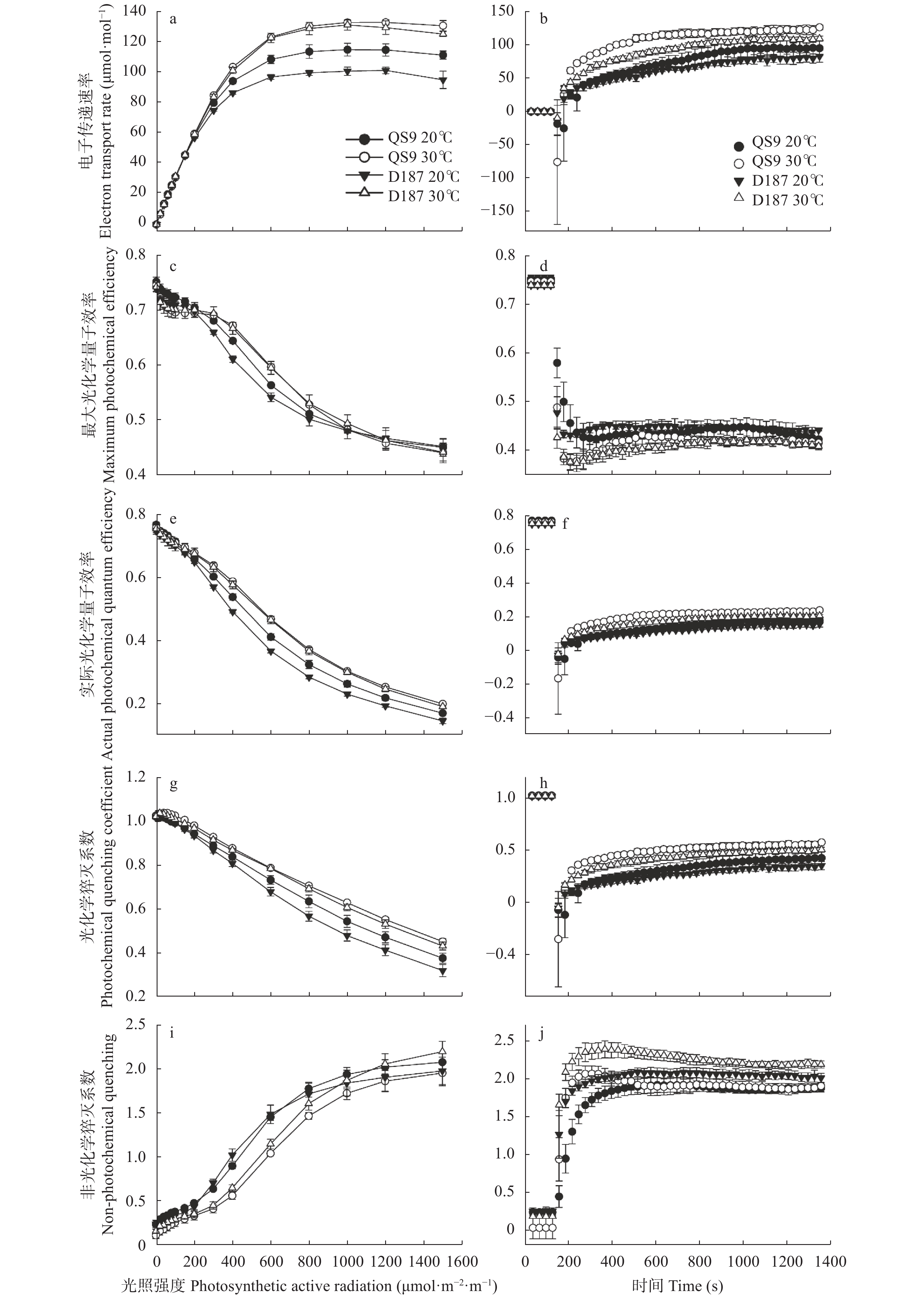
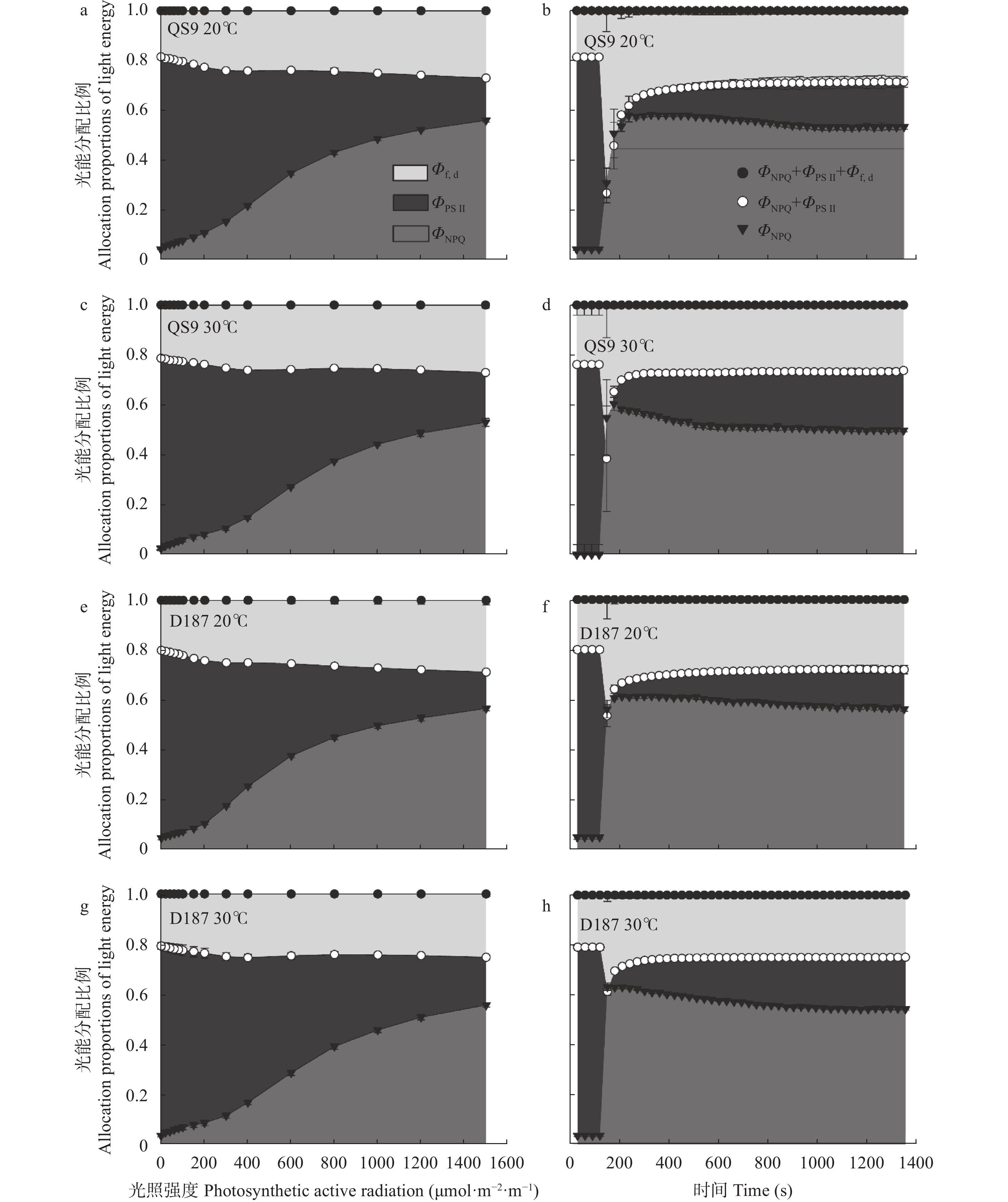
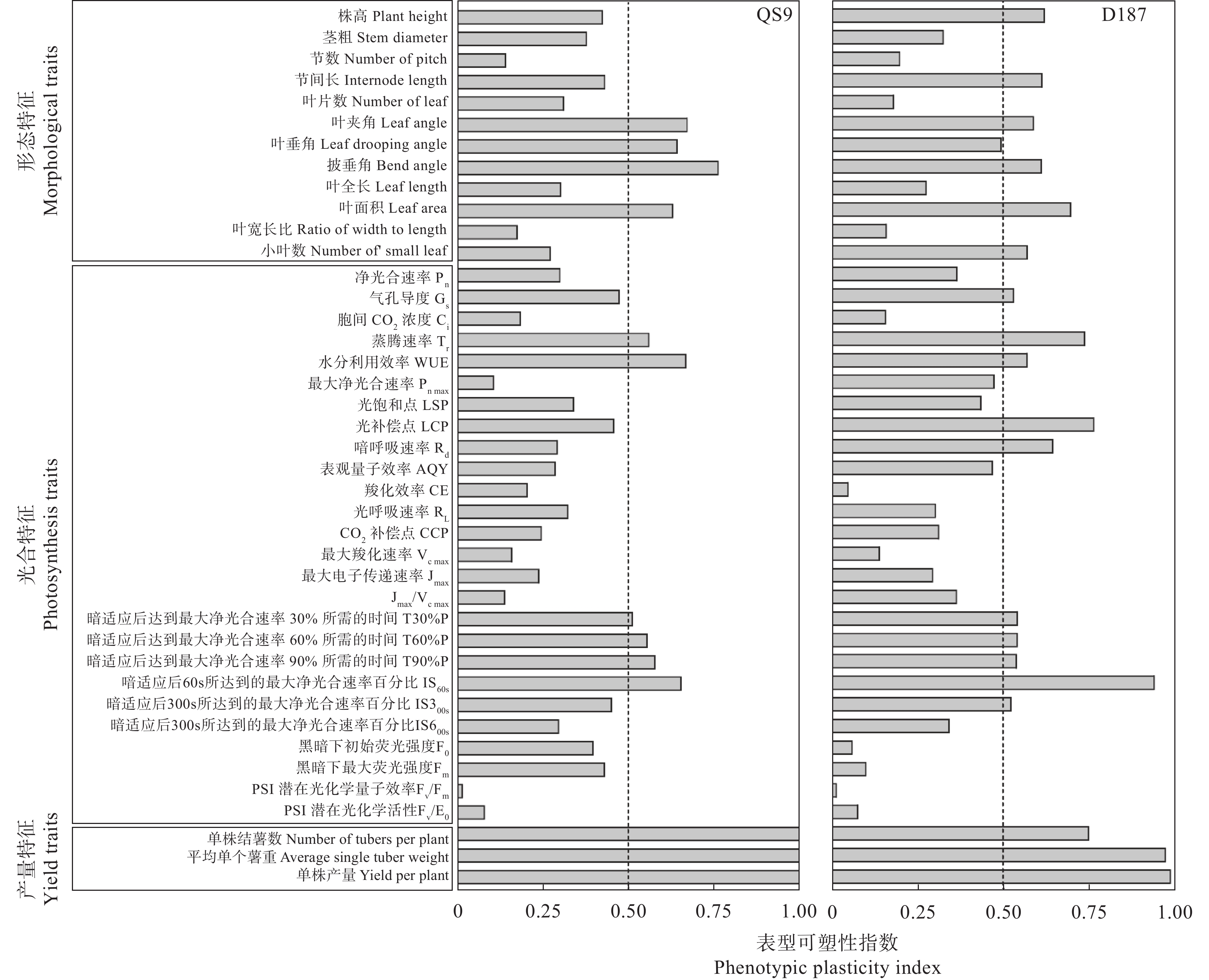
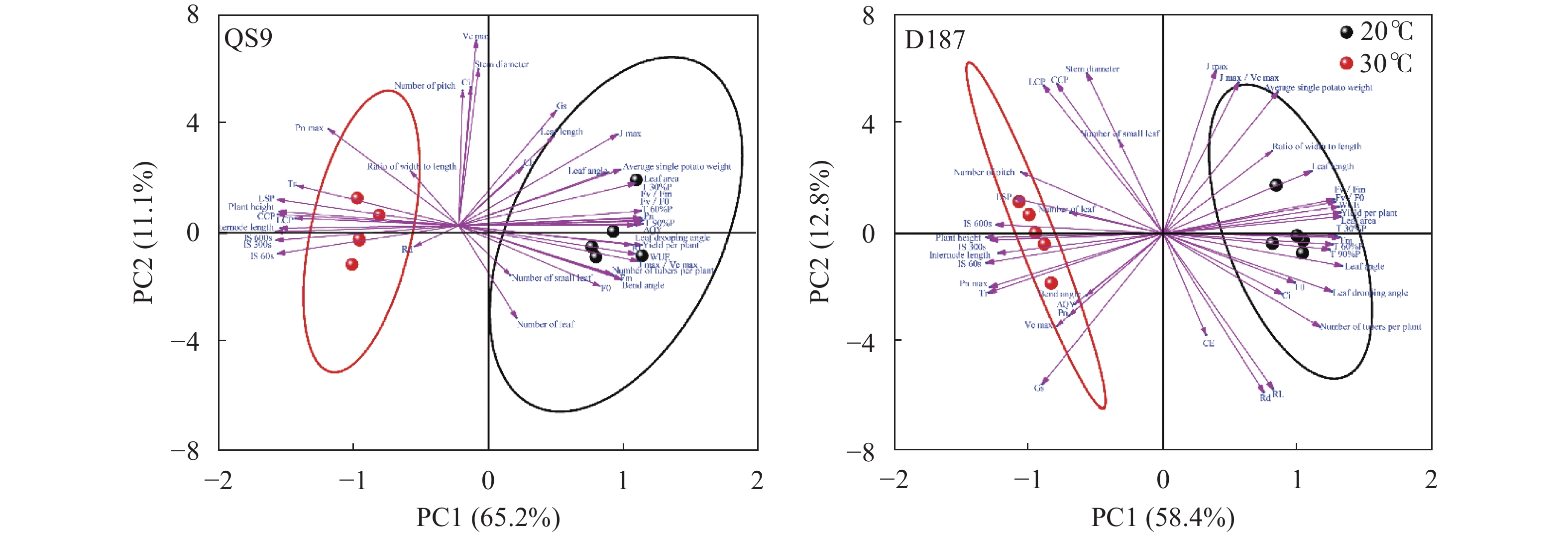
全球变暖对粮食生产的负面影响日益受到关注, 马铃薯是重要的粮菜兼用作物, 对高温敏感。探究耐热和热敏感型马铃薯资源在响应高温胁迫时的生理差异, 可为深入研究马铃薯耐热机制提供理论依据。本研究以耐热型品系‘滇187’(D187)和热敏感型品种‘青薯9号’(QS9)为材料, 在30 ℃高温胁迫处理2周后, 分析2个马铃薯材料在块茎形成期的植株形态和光合作用差异。在植株形态方面, 高温使马铃薯植株株高和节间长显著(P<0.01)增加, 叶片直立、叶片长度和面积缩小, 株型更为紧凑; 与QS9相比, D187叶片数目和披垂角更为稳定。高温胁迫下马铃薯植株水分散失加快, 水分利用率降低, 对CO2吸收和低浓度CO2利用能力减弱, 呼吸作用消耗增加, 1,5-二磷酸核酮糖(RuBP)的再生能力减弱, 黑暗下的叶绿素荧光参数降低, 光下叶绿素荧光参数升高, 对有限强光的利用能力增强。高温胁迫下, D187叶片具有更高的净光合速率、水分利用效率、最大净光合速率、表观量子效率、羧化效率、最大羧化速率、最大电子传递速率, 更低的光补偿点、暗呼吸速率, 说明D187光合能力更强、弱光利用率更高、呼吸消耗更低、碳同化能力更强。D187的形态和光合作用指标中, 可塑性指数大于0.5的参数均多于QS9, 平均可塑性指数(0.448)高于QS9 (0.418), 说明耐热型马铃薯能够更好地通过调节植株形态和光合作用来适应高温环境。


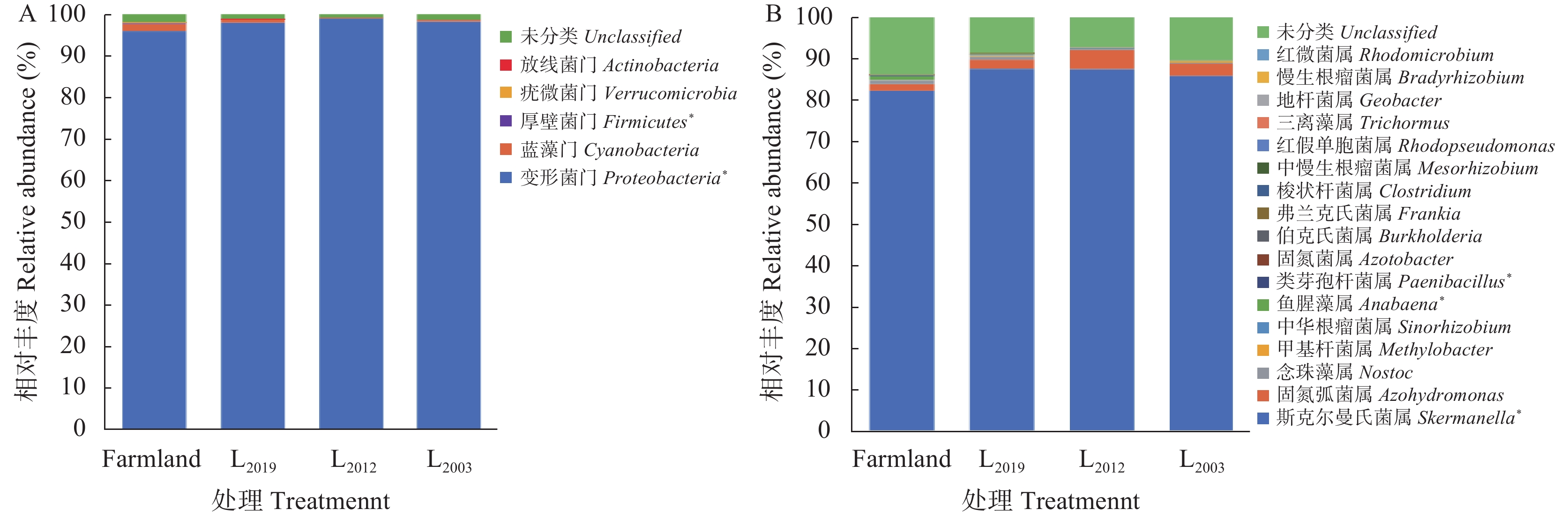
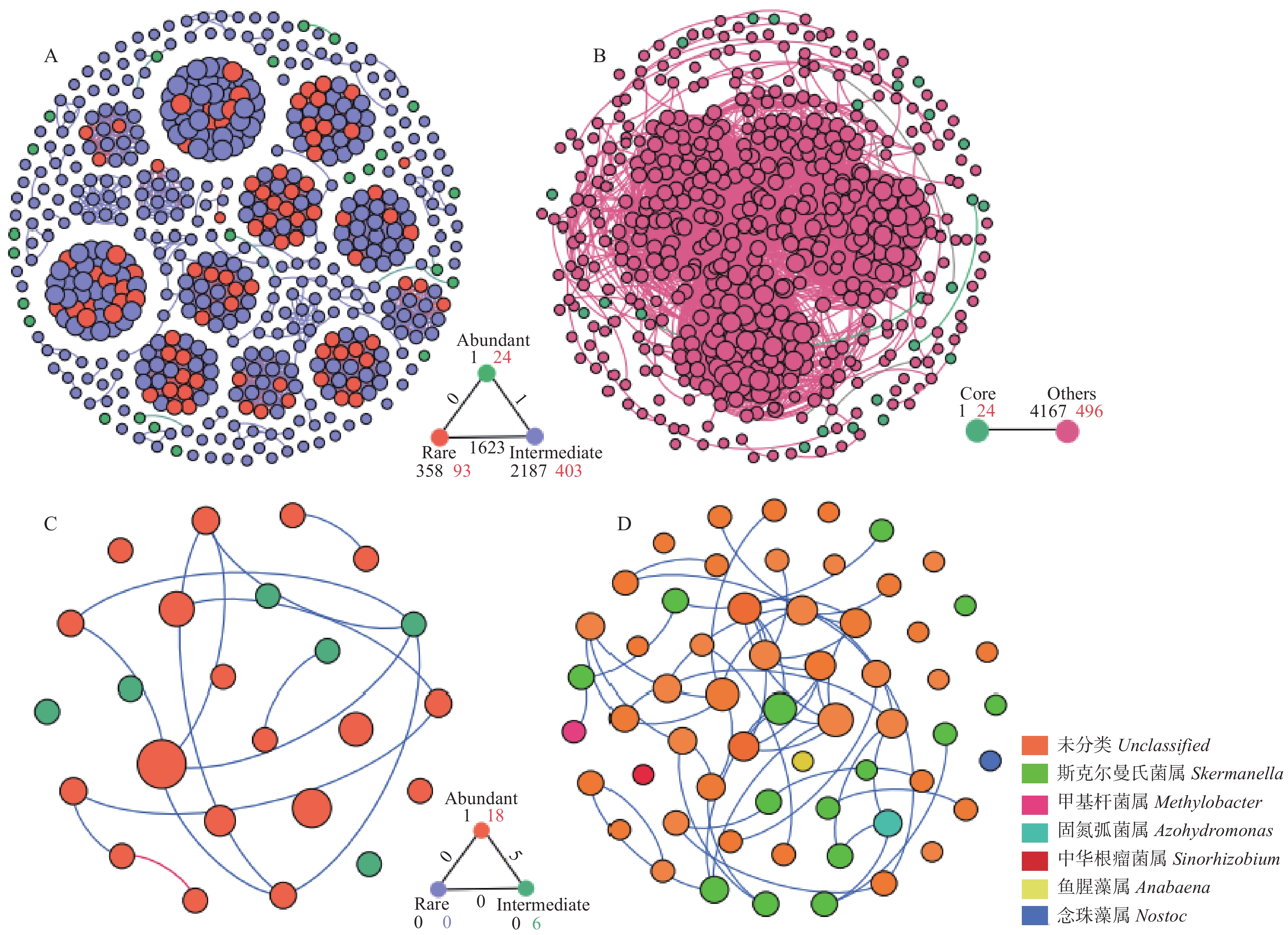
生物固氮是紫花苜蓿(Medicago sativa)土壤氮素的重要来源, 固氮微生物数量及其群落结构变化对土壤氮素供应和肥力维持起着重要作用。本研究采用Illumina MiSeq测序和荧光定量PCR技术, 探究了黄绵土区玉米农田和不同种植年限(2019年、2012年、2003年)紫花苜蓿地土壤nifH固氮基因丰度、nifH固氮微生物群落结构和多样性, 通过共现网络分析丰富和稀有固氮微生物的生态地位, 耦合土壤理化性质明确影响固氮微生物群落结构的主导因子。结果表明, 黄绵土固氮微生物nifH基因丰度为2.97×106~5.93×106 copies∙g−1 (干土), 且表现为苜蓿地显著高于玉米农田。土壤样品经测序共获得有效序列176 367条, 主要分布在5门、8纲、11目、15科、17属。门水平上, 变形菌门和蓝藻门为主要优势类群; 属水平以斯克尔曼氏菌属和固氮弧菌属为优势属。与玉米农田相比, 多年持续种植紫花苜蓿显著提高了斯克尔曼氏菌属的相对丰度, 但其随种植年限延长呈降低趋势。长期种植紫花苜蓿促生了固氮菌属、伯克氏菌属、弗兰克氏菌属、中慢生根瘤菌属、地杆菌属和慢生根瘤菌属等生理类群, 同时也使得梭状杆菌属、红假单胞菌属和三离藻属消亡。RDA分析发现, 固氮微生物不同种群对环境因子的响应并不一致, 具有各自的生态位, 但土壤全磷是影响土壤固氮微生物群落结构的主导环境因子, 其次是有机碳和硝态氮。分子生态网络分析表明固氮菌生态网络中丰富类群占据生态系统核心地位, 且物种间均为协同合作关系, 群落结构相对稳定, 对环境变化具有较强的适应能力。综上, 黄土高原半干旱区种植紫花苜蓿显著提高了土壤nifH基因丰度, 改变了固氮微生物nifH群落结构, 该结果可为黄绵土固氮微生物多样性研究和紫花苜蓿适宜种植年限的确定提供基础数据和理论依据。
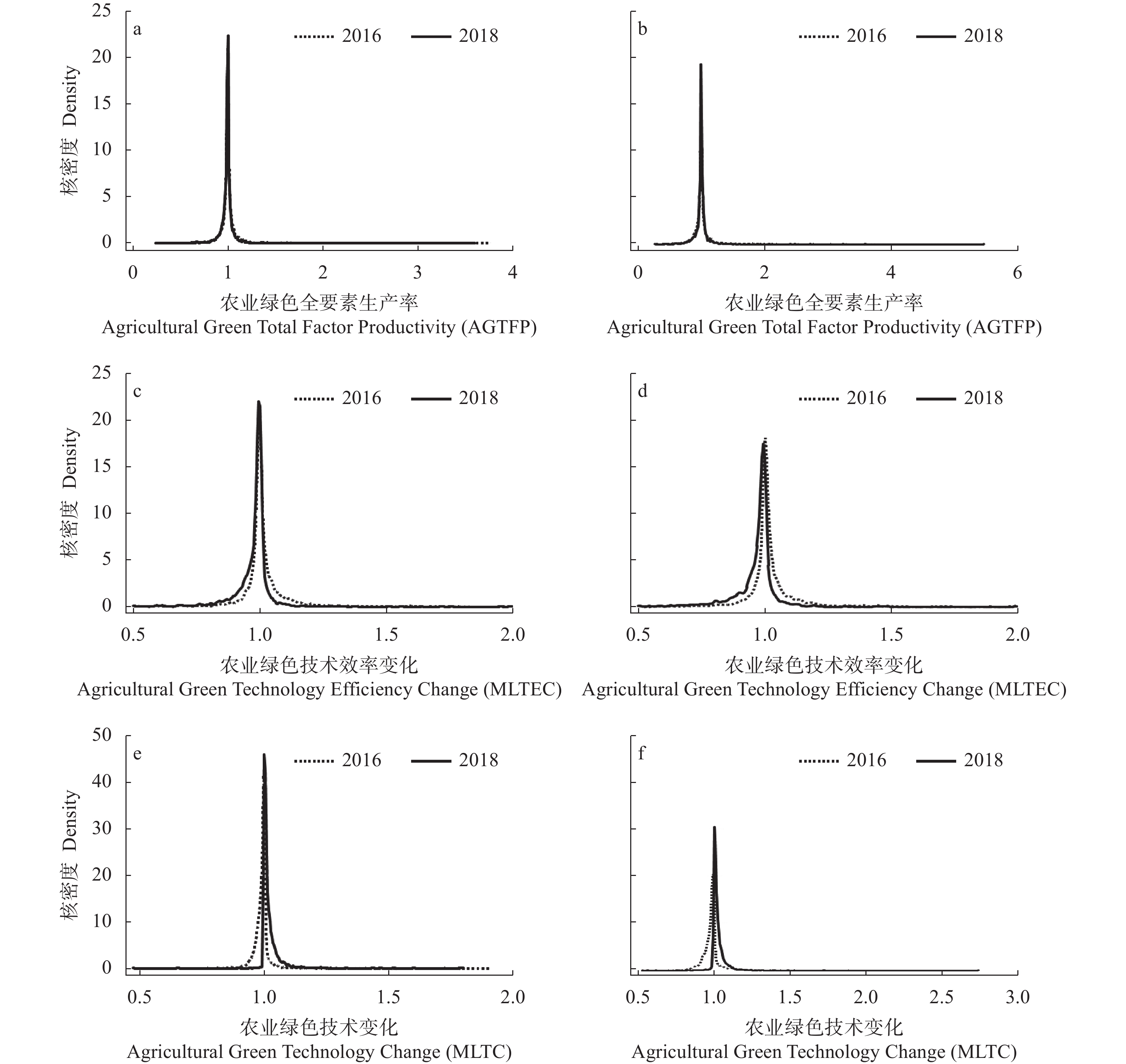
提升农业绿色全要素生产率, 加快农业绿色转型是全面建设社会主义现代化强国的必然选择。研究以中国家庭追踪调查(China Family Panel Studies, CFPS)的全国性大容量样本农户数据为蓝本, 在微观测度方法比较分析的基础上, 基于技术优化的Malmquist-Luenberger指数为基准, 测度分析了农户层农业绿色全要素生产率的状况, 并进一步选用核密度估计法和Dagum基尼系数法, 揭示呈现了微观样本农业绿色全要素生产率的动态演变规律及其区域差异特征。主要研究发现如下: 1)技术优化的Malmquist-Luenberger指数测度显示, 2014、2016和2018年3期样本农户的农业绿色全要素生产率均值为1.0030, 总体发展态势良好; 农业绿色技术变化、绿色技术效率变化的共同作用是驱动农户层面农业绿色发展变化的主要引致因素, 且后者的影响程度远大于前者; 农户资源配置、管理模式及组织方式的改善优化, 在现阶段是农户发展绿色农业的提升关键, 其影响相对高于农户农业生产技术的革新。2)通过核密度估算发现, 2016和2018年样本农户的绿色全要素生产率集中度较高, 农业绿色技术效率并未出现两级分化, 但农业绿色技术进步呈现上升趋势。3) Dagum基尼系数法结果表明, 农户层面农业绿色全要素生产率的区域差距不断缩小, 区域差距的降幅达22.32%, 超变密度是引致主因; 在区域内差距上, 东、西、中部地区内部, 农户的绿色农业差距依次递减; 在区域间差距上, 东西、东中、中西部间差距不断缩小、协同性不断增强, 但差距易受到环境因素影响。
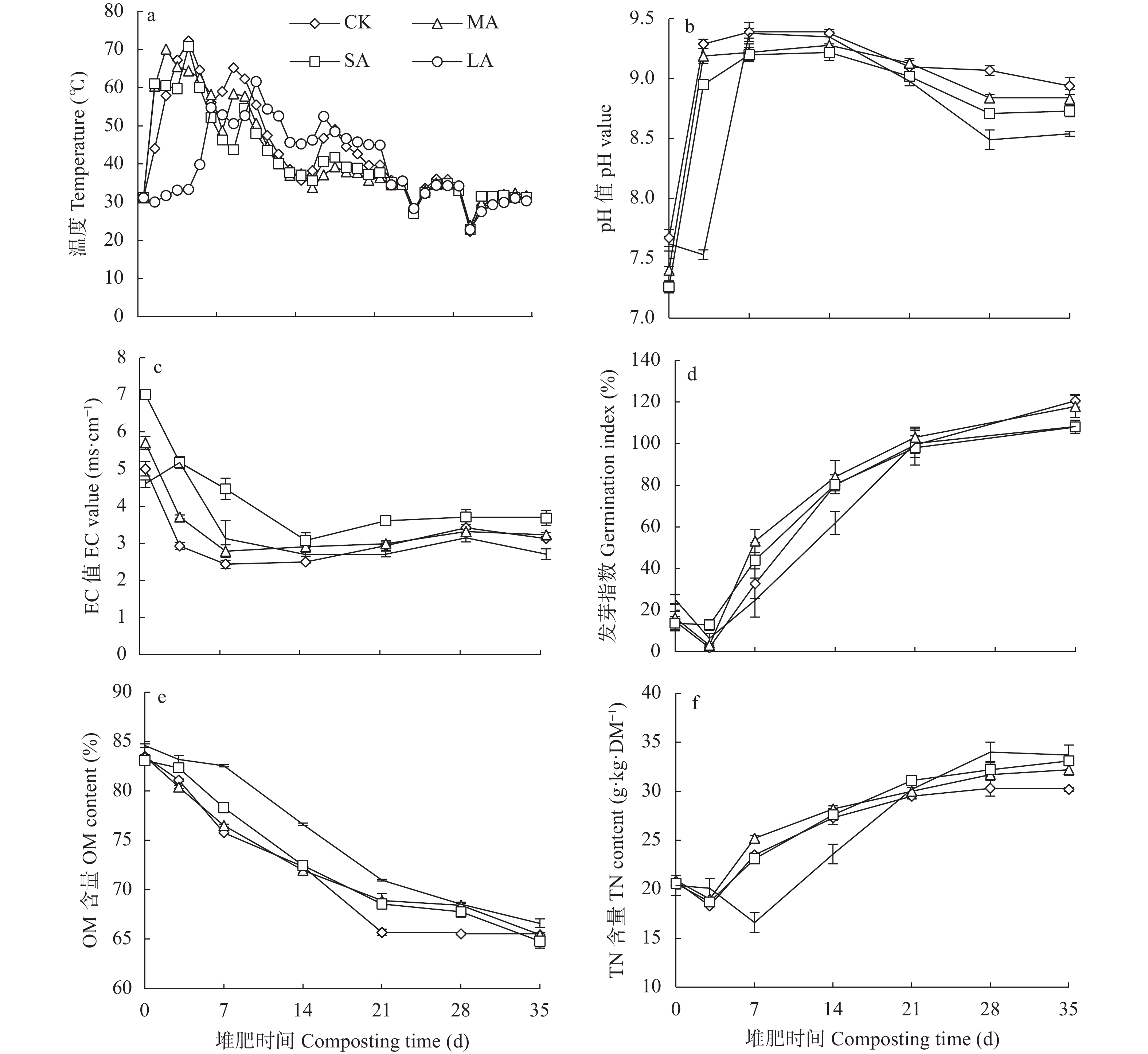
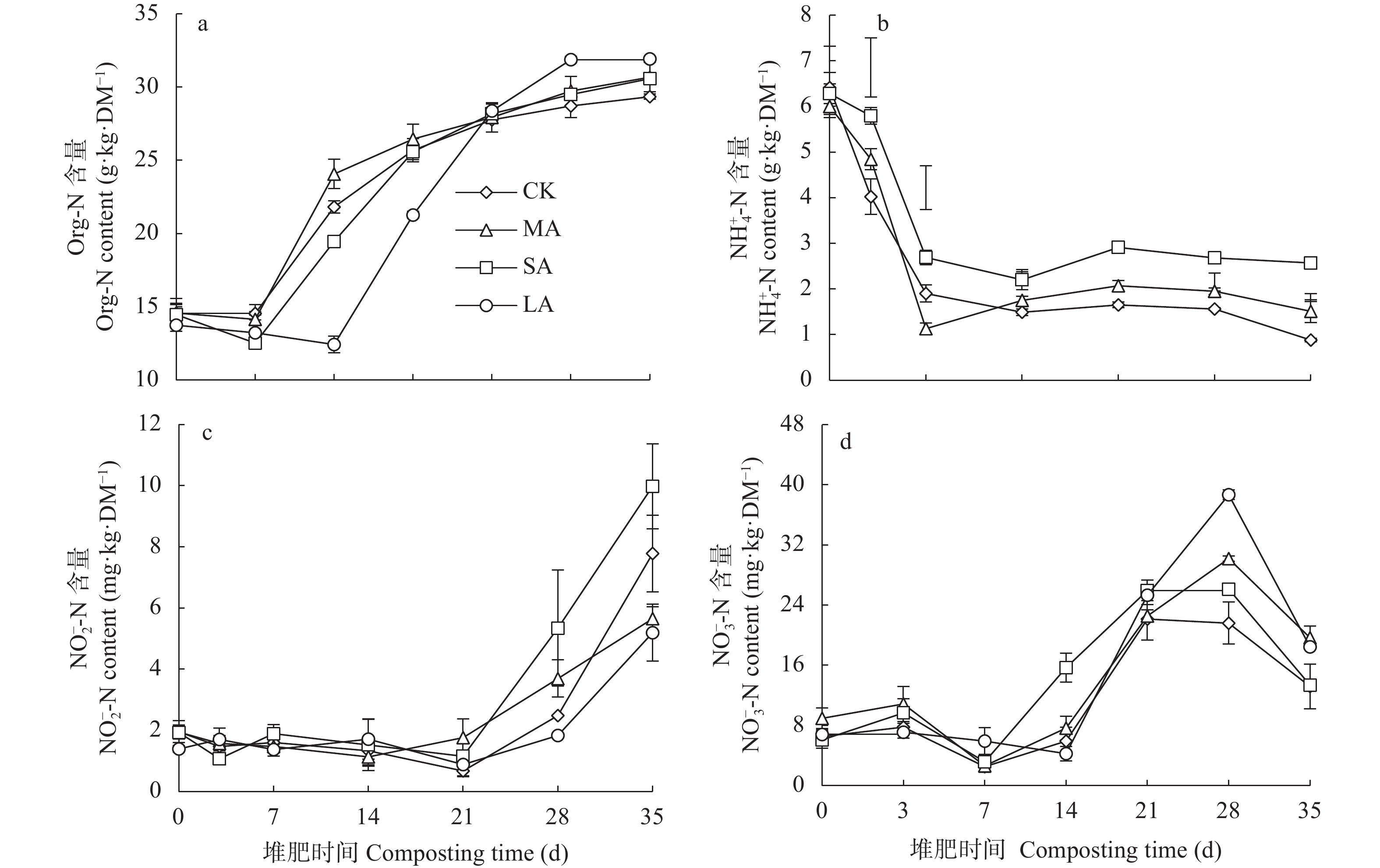
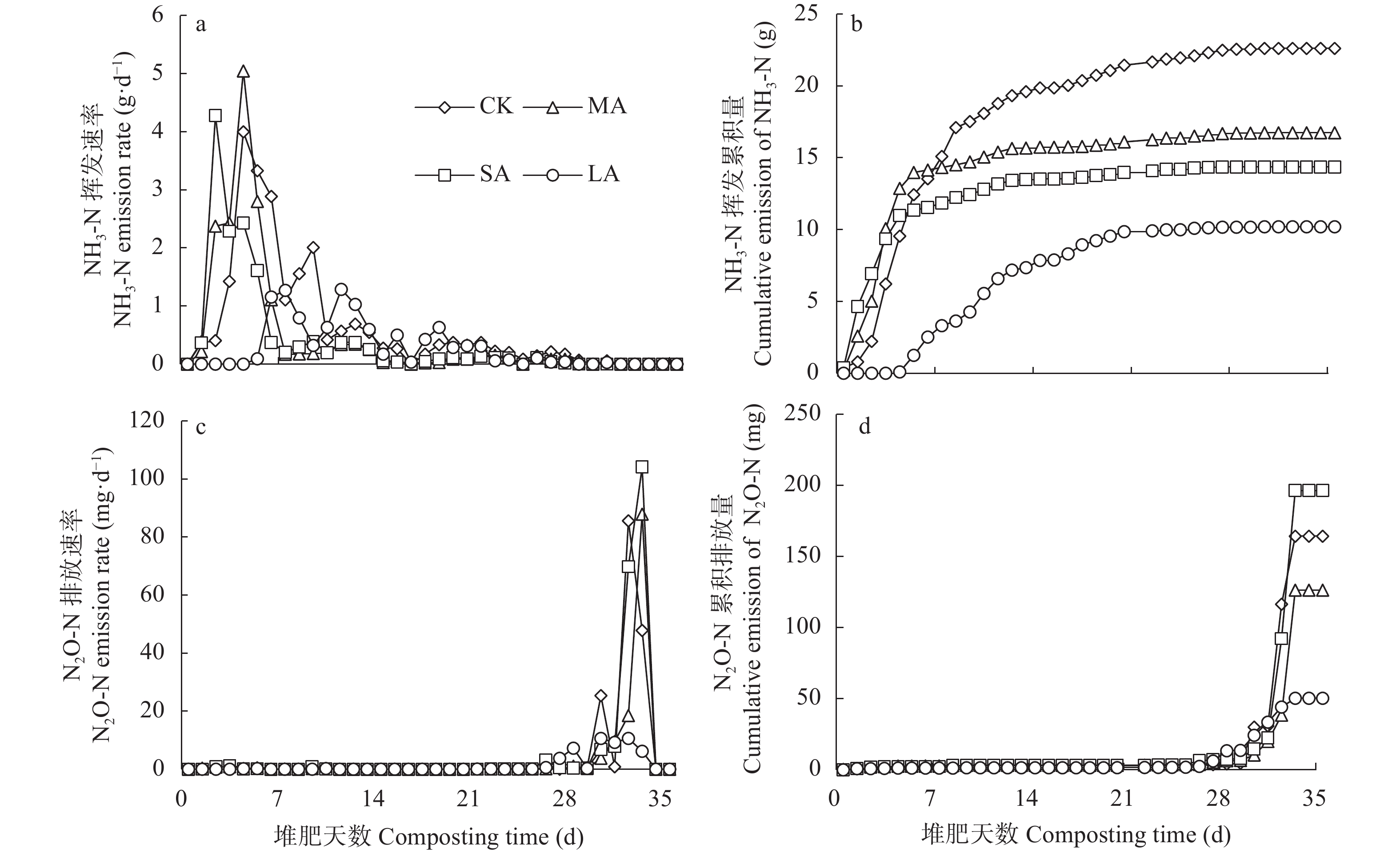
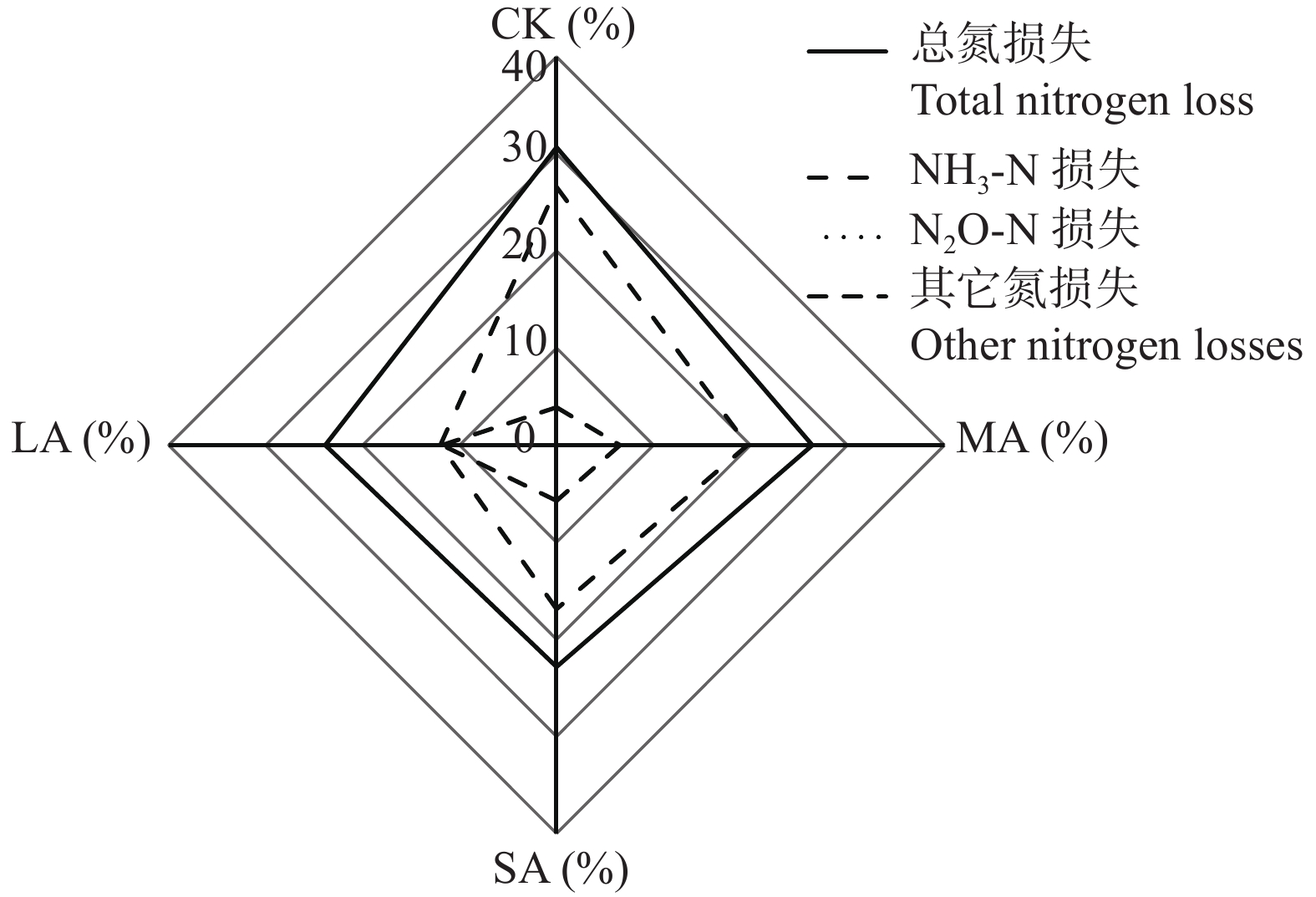
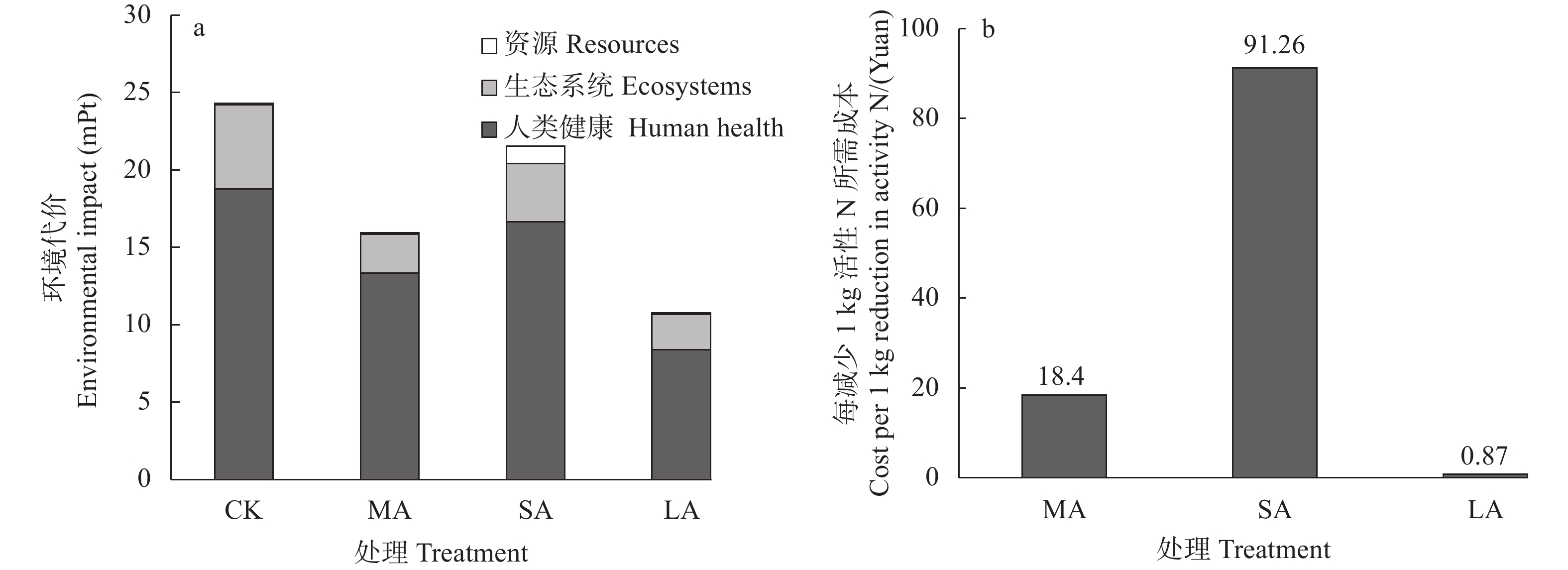
酸化是减少堆肥过程中氮素损失的有效手段, 而传统无机酸酸化具有成本高、二次污染严重等缺点, 探究新型酸化工艺对减少堆肥过程中的养分流失和环境污染具有重要意义。本研究以食品残渣(果渣、豆渣)为基质, 通过乳酸菌厌氧发酵制备了一种富含乳酸(70 mmol∙L−1)和乳酸菌(106 cfu∙mL−1)的酸性调理剂, 用于农业废弃物(猪粪、小麦秸秆)酸化堆肥试验,设置两种新型酸化方式处理: 添加30%酸性调理剂处理(MA)和添加3%酸性调理剂的厌氧自酸化处理(LA), 同时以不加酸处理(CK) 添加硫酸处理(SA)作为对照。通过分析堆肥过程中理化性质和氮素形态等变化发现, 3种酸化方式的堆肥产品均达到腐熟标准(发芽指数>80), 其中MA处理的腐熟程度最优(发芽指数=117.8%); MA、SA、LA处理的总氮损失较CK分别显著降低14.0%、25.6%、22.2% (P<0.05), 其中NH3挥发量较CK分别显著减少26.0%、36.5%、54.9% (P<0.05); 酸化处理提高了NH4+含量, 促进了硝化过程, 又间接增强了反硝化过程, MA、LA处理显著减少23.1%、69.4%的N2O排放(P<0.05), 而SA处理抑制了N2O的还原, 增加18.3%的N2O的排放; 同时MA、SA、LA处理总环境代价相较于CK分别显著降低34.5%、11.0%、55.9% (P<0.05), 且MA、LA每减少1 kg活性氮排放分别需要18.4元、0.9元, 远低于SA处理(91.26元)。综上所述, MA、LA处理可作为降低堆肥过程中氮损失的可行方法, 本研究为堆肥酸化保氮技术提供了理论依据。
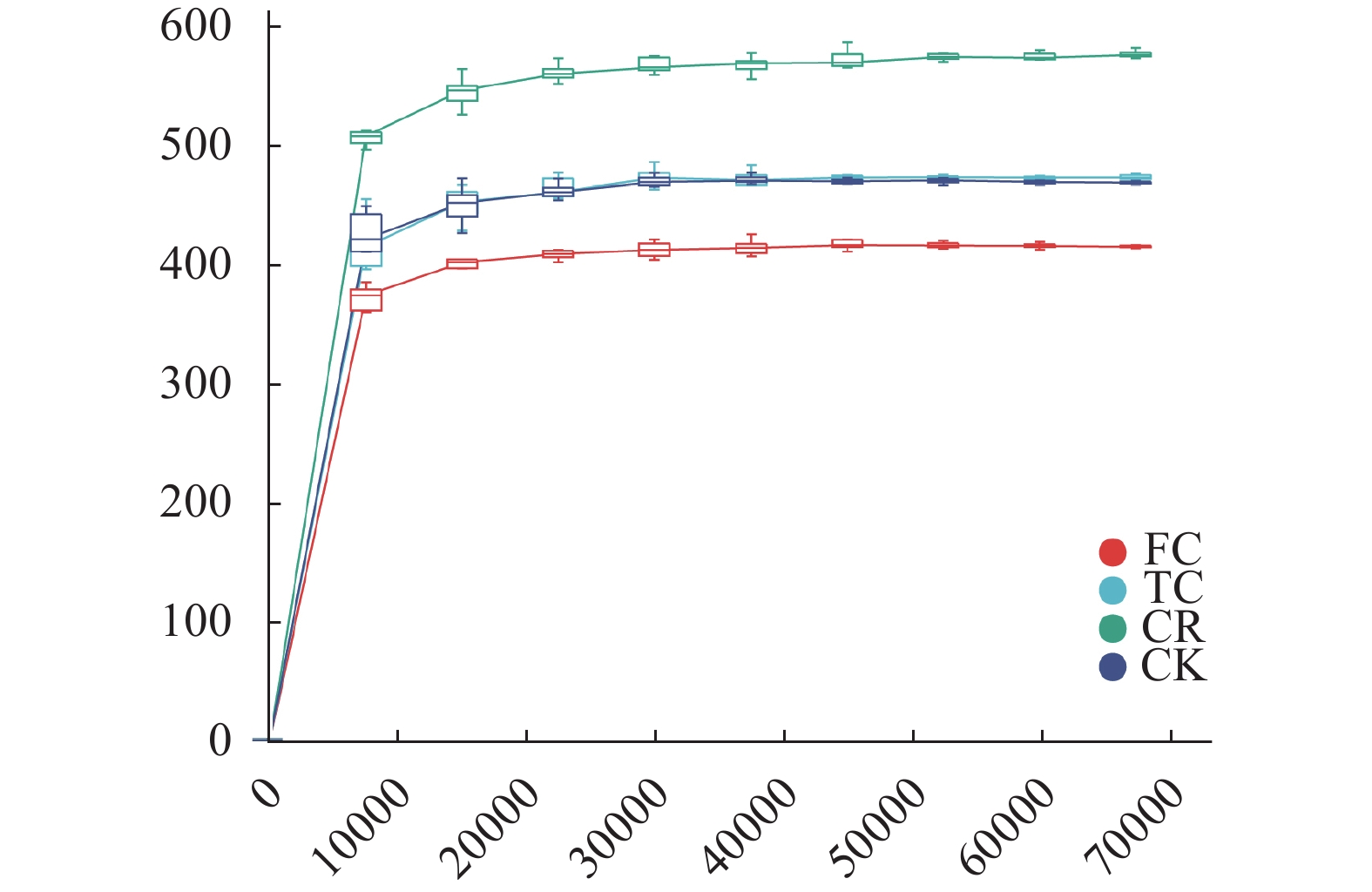
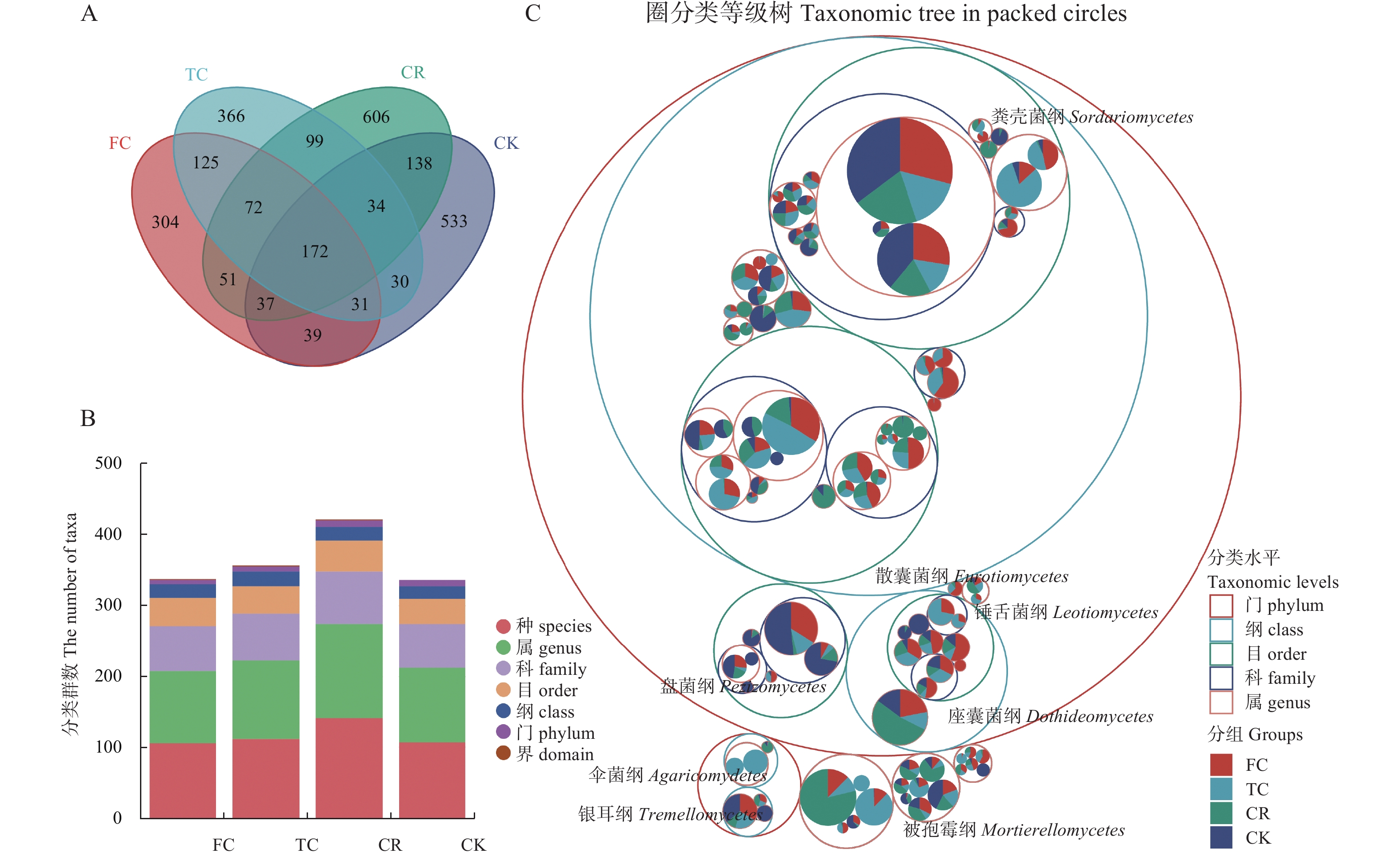
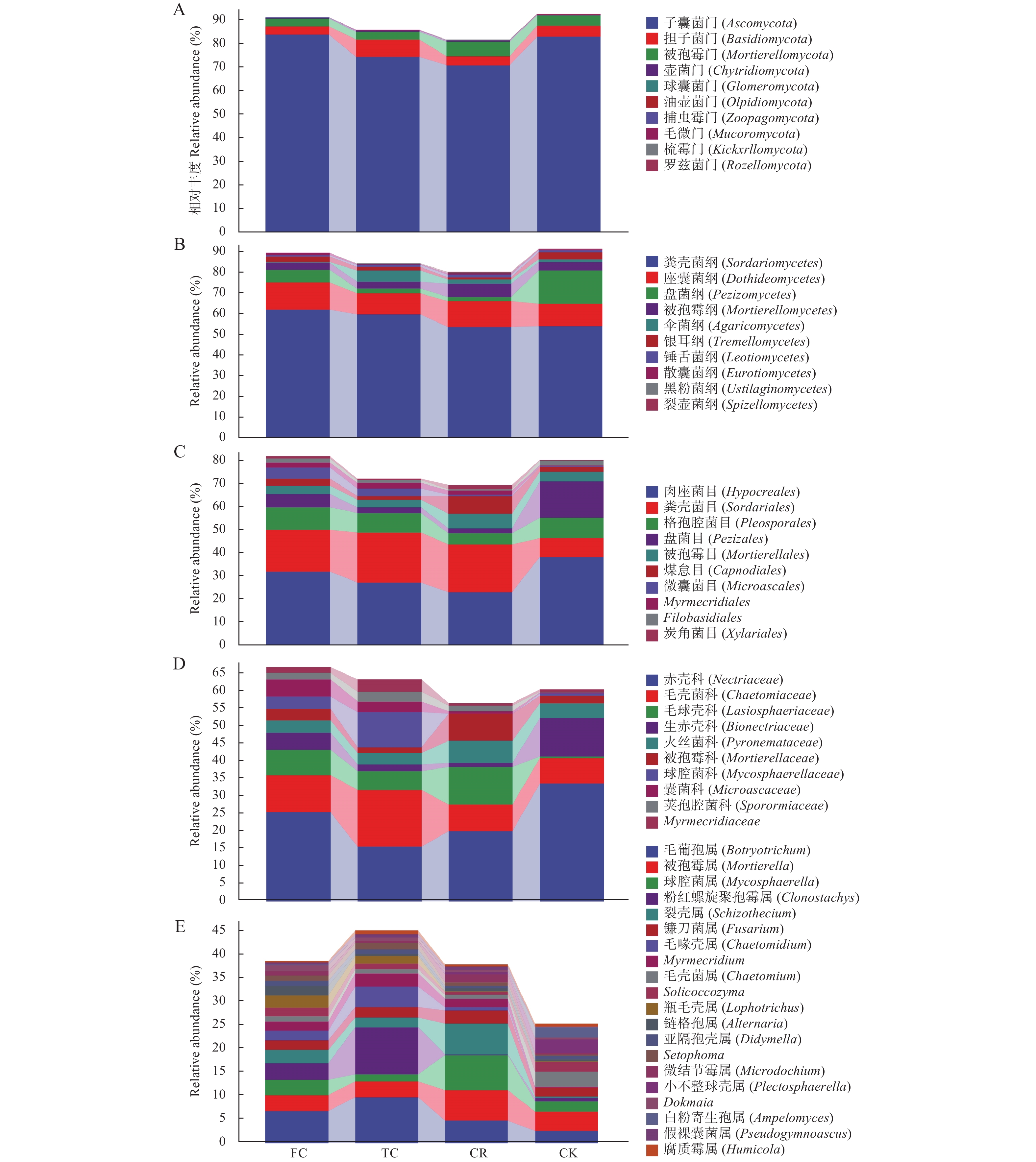
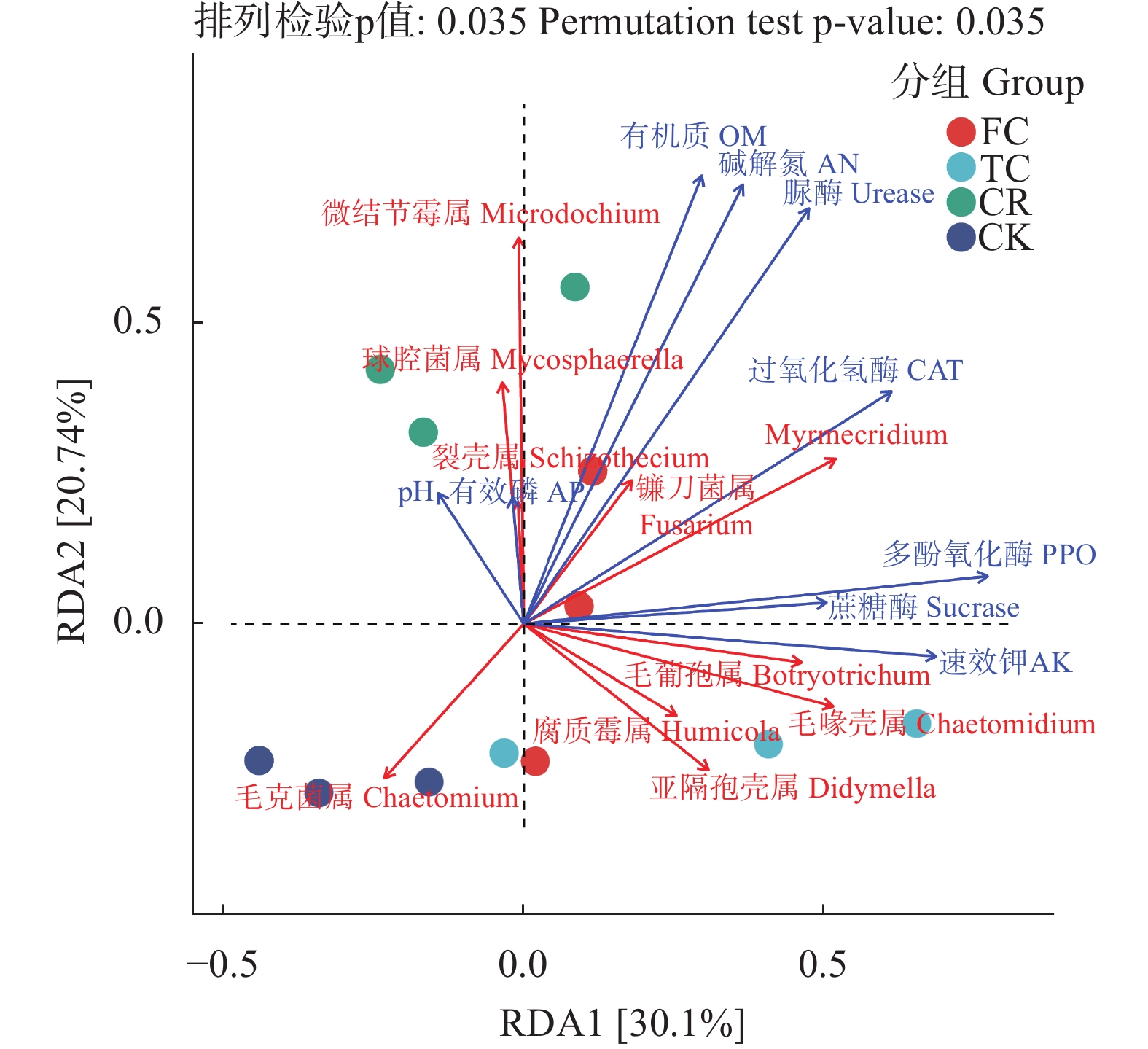
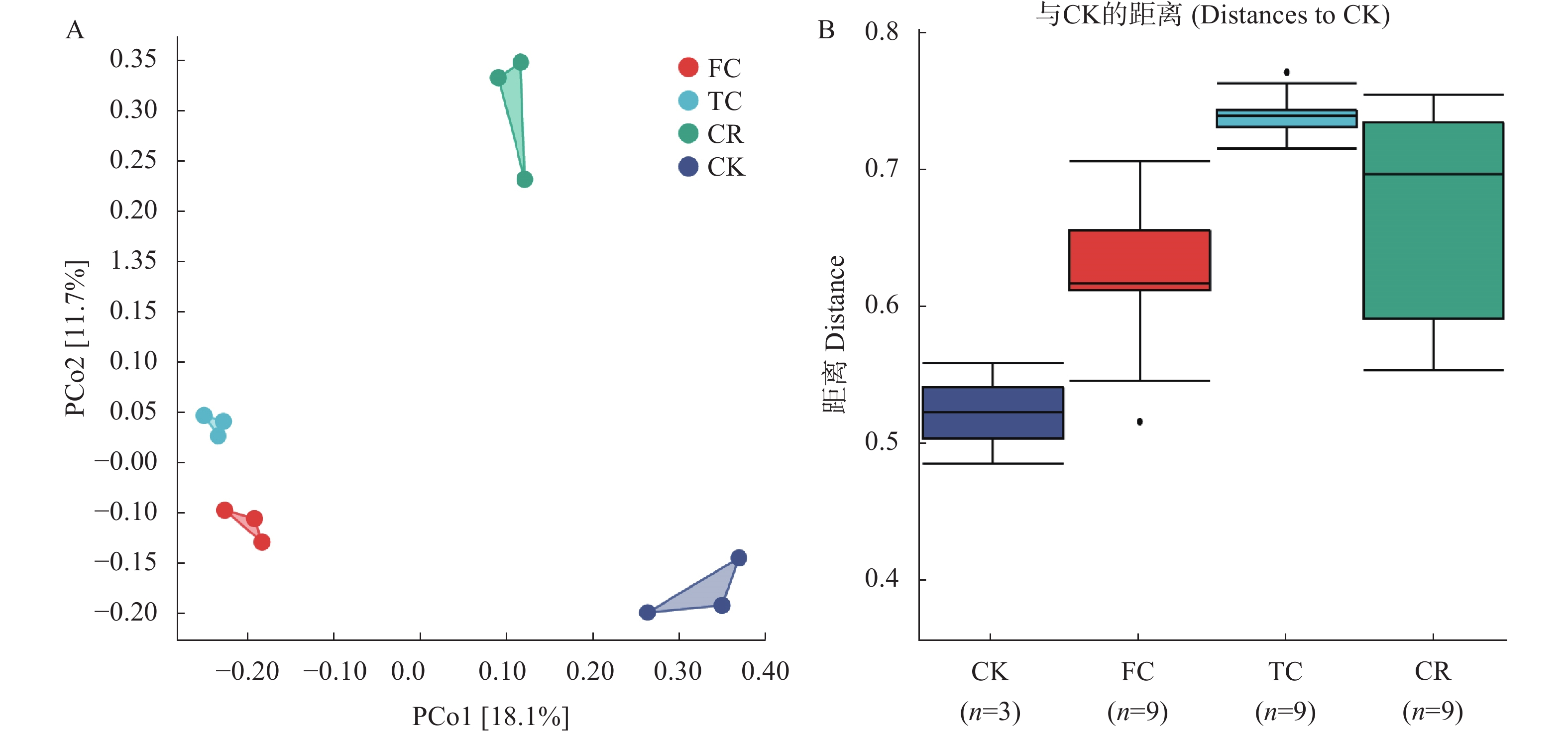
为了解谷子连作对土壤真菌群落结构的影响, 以撂荒地为对照, 以谷子-玉米轮作、谷子连作3年、连作5年根际土壤为研究对象, 采用真菌ITS高通量测序技术, 探究不同种植模式下谷子土壤真菌群落分布特征。结果表明: 不同种植模式下, 谷子根际土壤共检测到真菌10门24纲46目79科136属和146种。在门和纲水平上群体结构相对稳定, 谷子田土壤优势门主要包括子囊菌门和担子菌门, 优势纲为粪壳菌纲、座囊菌纲和盘菌纲, 在目水平谷子根际土壤粪壳菌目相对丰度是撂荒地的2倍以上, 在科水平和属水平轮作土壤被孢霉、球腔菌相对丰度高于连作土壤, 链格孢菌、亚隔孢壳菌和粉红螺旋聚孢霉菌相对丰度低于连作土壤。Alpha多样性分析显示, 谷子-玉米轮作与谷子连作根际土壤真菌丰度差异达显著水平(P<0.05), 轮作土壤真菌丰度最高。Beta多样性分析显示连作3年和连作5年根际土壤真菌结构相似, 与撂荒地以及轮作根际土壤真菌结构存在差异, 表明不同种植模式谷子根际土壤真菌群落结构发生了改变。相关性分析显示, 碱解氮与有机质呈极显著正相关(P<0.01), 与有效磷、脲酶活性呈显著相关(P<0.05), 多酚氧化酶活性与速效钾呈显著正相关(P<0.05)Chao1指数、Observed species指数与多酚氧化酶活性呈极显著正相关(P<0.01)。冗余分析(RDA)表明, CK受毛壳菌影响, CR受球腔菌属和微结节霉属的影响, TC和FC受毛葡孢属、毛喙壳属、亚隔孢壳属等影响。LEfSe分析确定了谷子根际土壤特定标志物, 轮作根际土壤的标志物包含被孢霉属和球腔菌属, 连作3年根际土壤标志物包含毛葡孢属、亚隔孢壳属和粉红螺旋聚孢霉属, 连作5年根际土壤标志物包含链格孢菌属和亚隔孢壳属。因此, 谷子-玉米轮作与谷子连作相比, 土壤真菌群落结构差异较大, 轮作土壤腐生菌较多, 连作土壤病原菌较多, 为谷子连作障碍研究提供了有用信息。

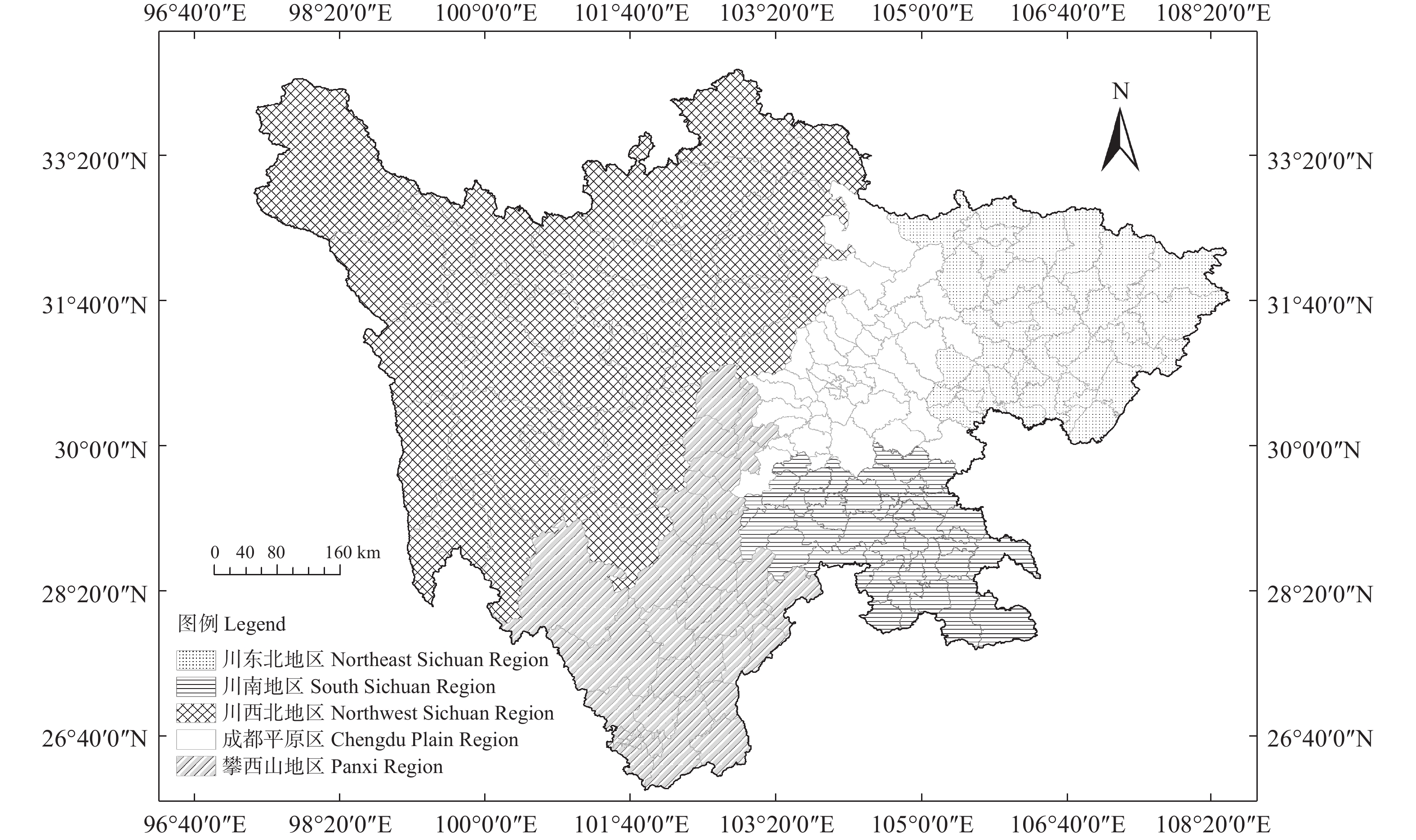
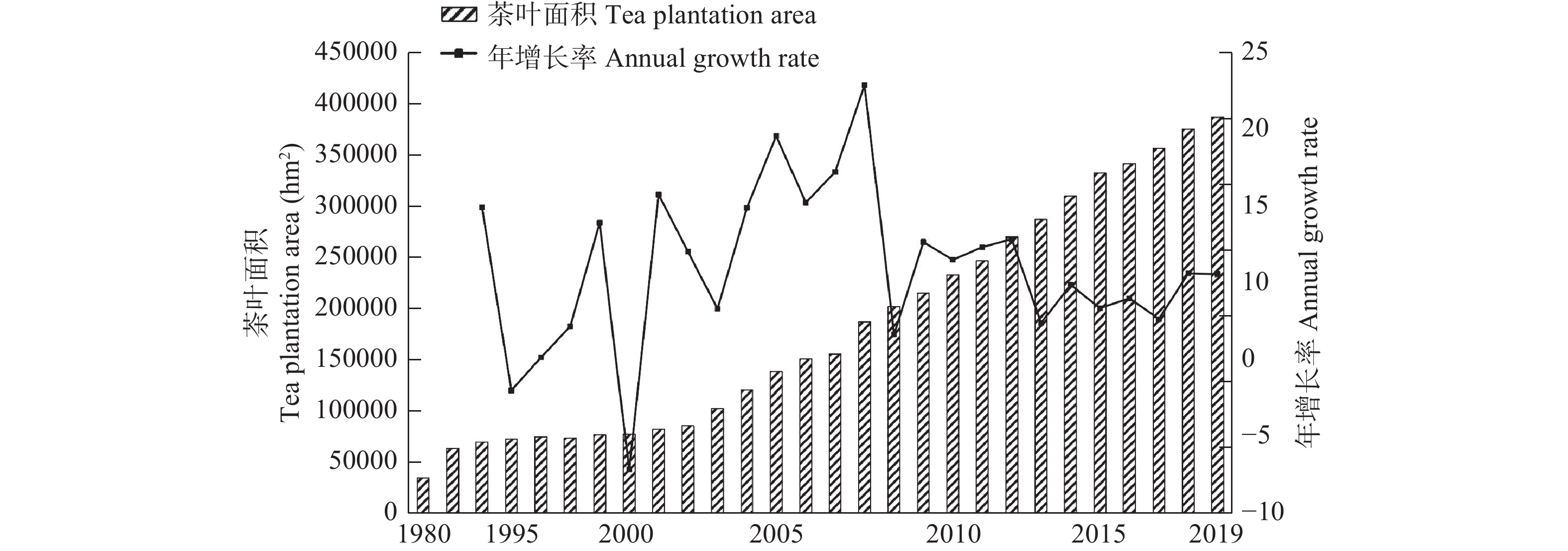
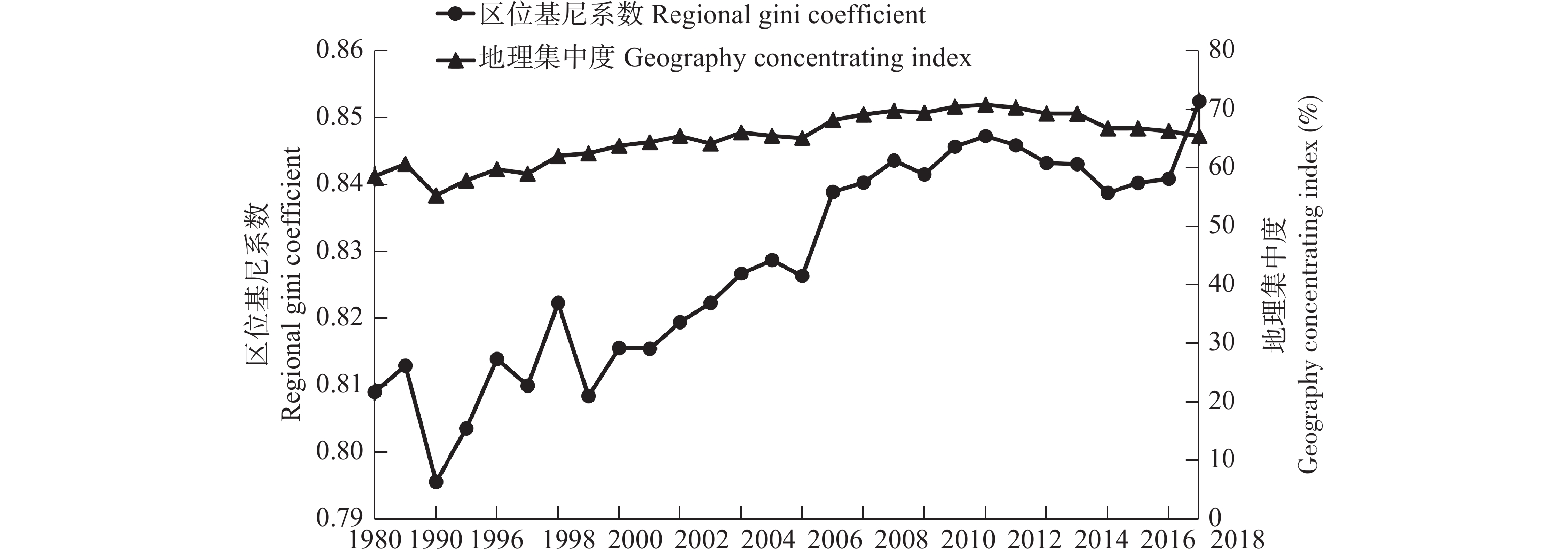
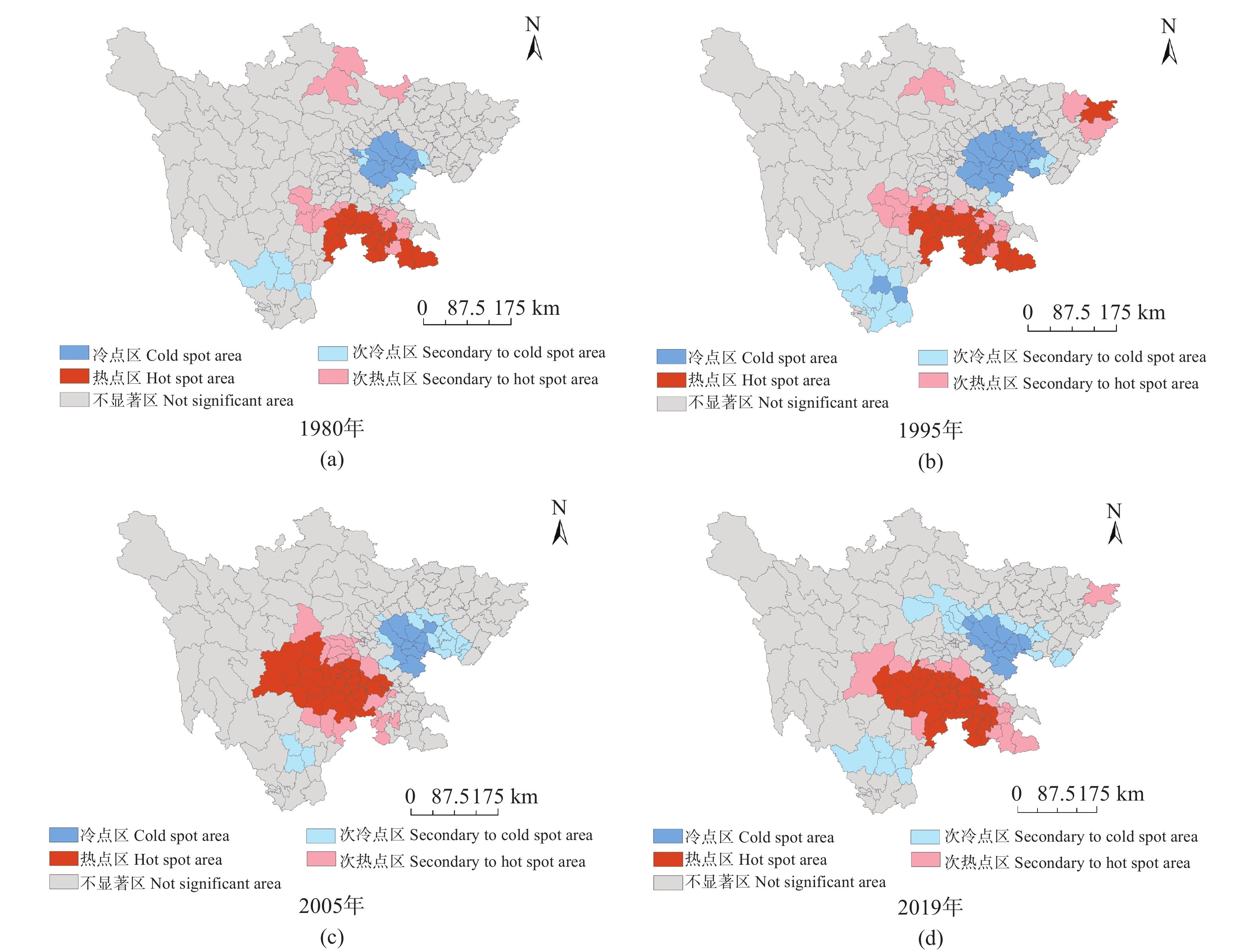
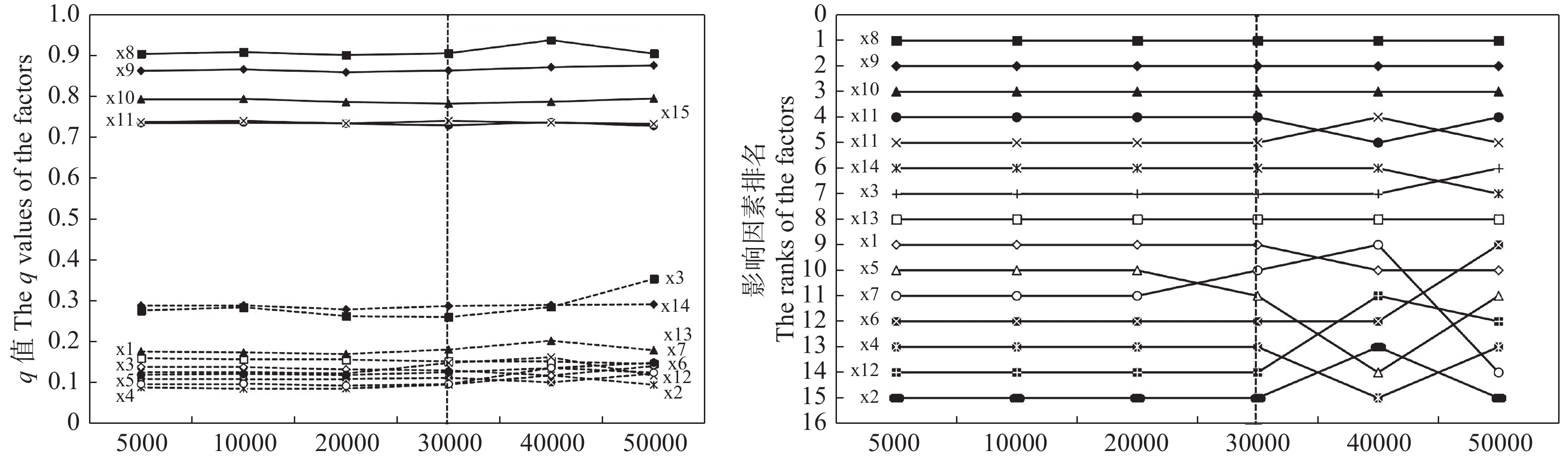
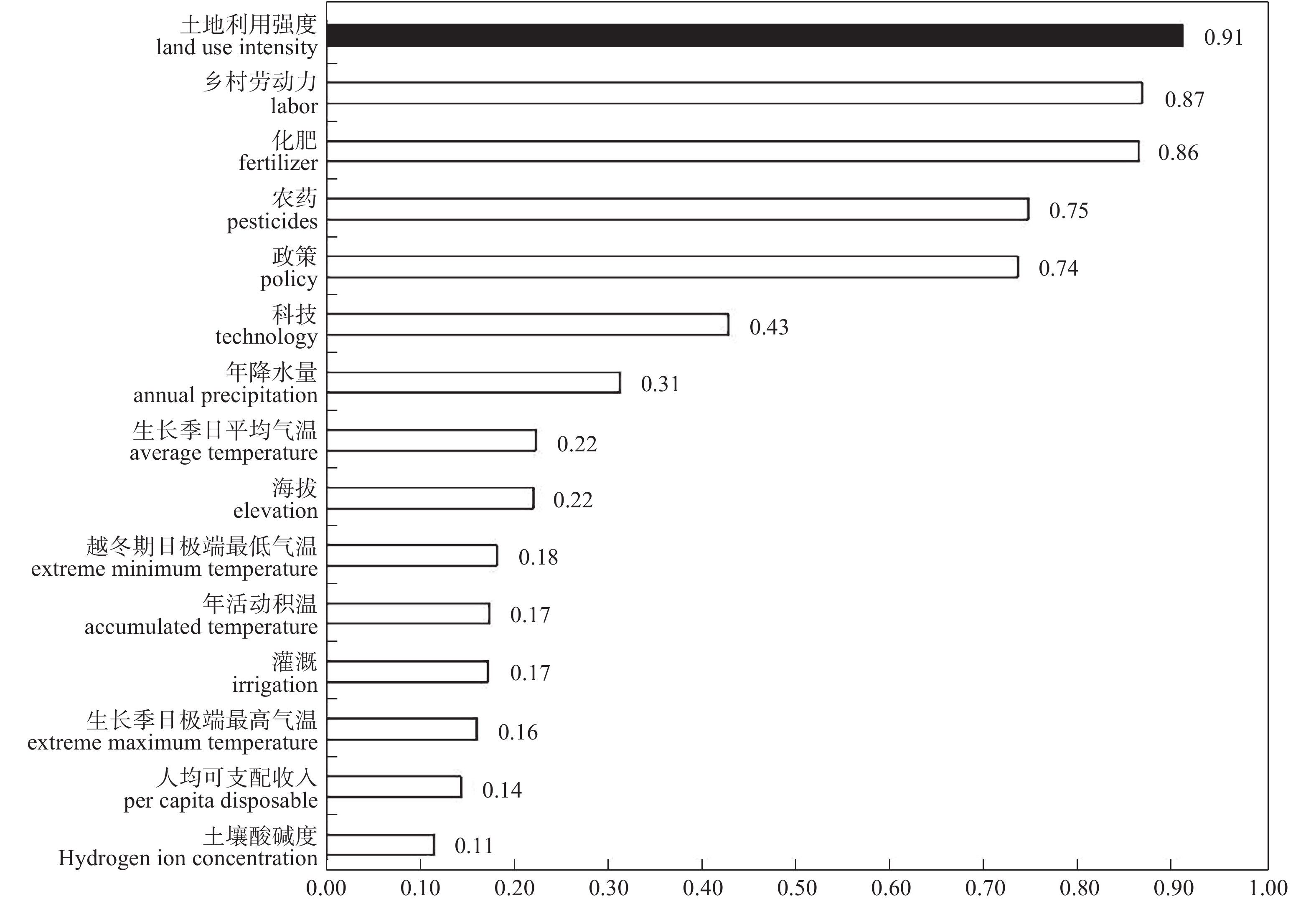
茶产业时空格局形成和演变是自然因素和人类活动共同作用的结果, 理解茶产业时空格局变化过程, 揭示不同自然—社会经济驱动因子对茶产业时空格局演变的作用机制, 对区域茶叶种植结构调整具有重要意义。本文基于1980—2019年四川省县区尺度茶叶生产统计年鉴数据, 运用产业集中度、探索性空间数据分析和产业重心模型分析了四川省茶产业时空格局演化过程, 对研究区内海拔、土壤酸碱度、年降水量、年活动积温、生长季日平均气温、越冬期日极端最低气温、生长季日极端最高气温等自然因素, 土地利用强度、乡村劳动力、化肥、农药、灌溉等生产要素以及人均可支配收入、科技、政策等社会经济要素进行离散分层并确定最优尺度单元, 基于地理探测器探讨了各驱动因子对四川省茶产业分布的解释力以及交互作用。结果表明: 从时间上看, 四川省茶产业规模总体呈上升趋势, 区位基尼系数均大于0.5, 空间特征呈现出高度集聚, 且集聚程度随时间波动上升。从空间上看, 全局莫兰指数均大于0, 县域尺度上表现出明显的空间集聚, 且相邻县域之间相互影响, 热点区主要分布在川南地区和成都平原区南部, 茶产业重心整体上向西迁移。可变面域问题会影响地理探测器建模结果, 对连续型因子离散化和空间单元尺度优化, 得到最优参数。单个因子对茶产业空间影响程度排前三的是, 土地利用强度(0.91)、乡村劳动力(0.87)、化肥(0.86); 影响因子相互作用主要表现为非线性增强和双因子增强, 生产与社会经济因子平均交互作用最大(0.8870), 四川省茶产业表现出生产要素驱动为主的空间格局。基于此, 本研究认为应重视: 1)关注生长季缺水, 突发性强降水以及低温冻害对茶树的影响; 2)加强“宜机采”茶园建设, 树立绿色茶园绿色发展概念; 3)提升良种普及率以及推广新技术, 保障用地、劳动力、化肥、农药等生产要素的稳定投入。
为探究纳米硅对低温下番茄根系构型及碳水化合物积累的调控机制, 以番茄品种‘中杂9号’为材料, 通过基质盆栽试验, 研究了施用纳米硅对低温下番茄幼苗生物量、根系构型、光合能力以及非结构性碳水化合物含量的影响。结果表明: 1)低温下番茄幼苗生物量、总根长、根尖数、光合色素含量和净光合速率等显著下降(P<0.05), 可溶性糖、蔗糖和淀粉含量显著增加(P<0.05), 其中地上部鲜重、净光合速率和总根长分别降低48.60%、66.88%和65.49% (P<0.05)。2)施用纳米硅在常温和低温下均能显著提高番茄幼苗的生物量、根系活力、根尖数、分形维数、净光合速率和非结构性碳水化合物含量等, 其中低温下施用纳米硅番茄幼苗根分叉数、净光合速率和叶片可溶性糖含量分别提高35.25%、48.24%和75.69% (P<0.05)。由上可知, 低温严重制约了番茄的光合作用、根系的生长发育以及非结构性碳水化合物的积累, 根系构型参数偏向于不利于植物正常生长的方向变化, 施用纳米硅可通过促进光合色素合成、提高光合速率和根系活力、改善根系构型及提高非结构性碳水化合物积累来提高番茄抗冷性。
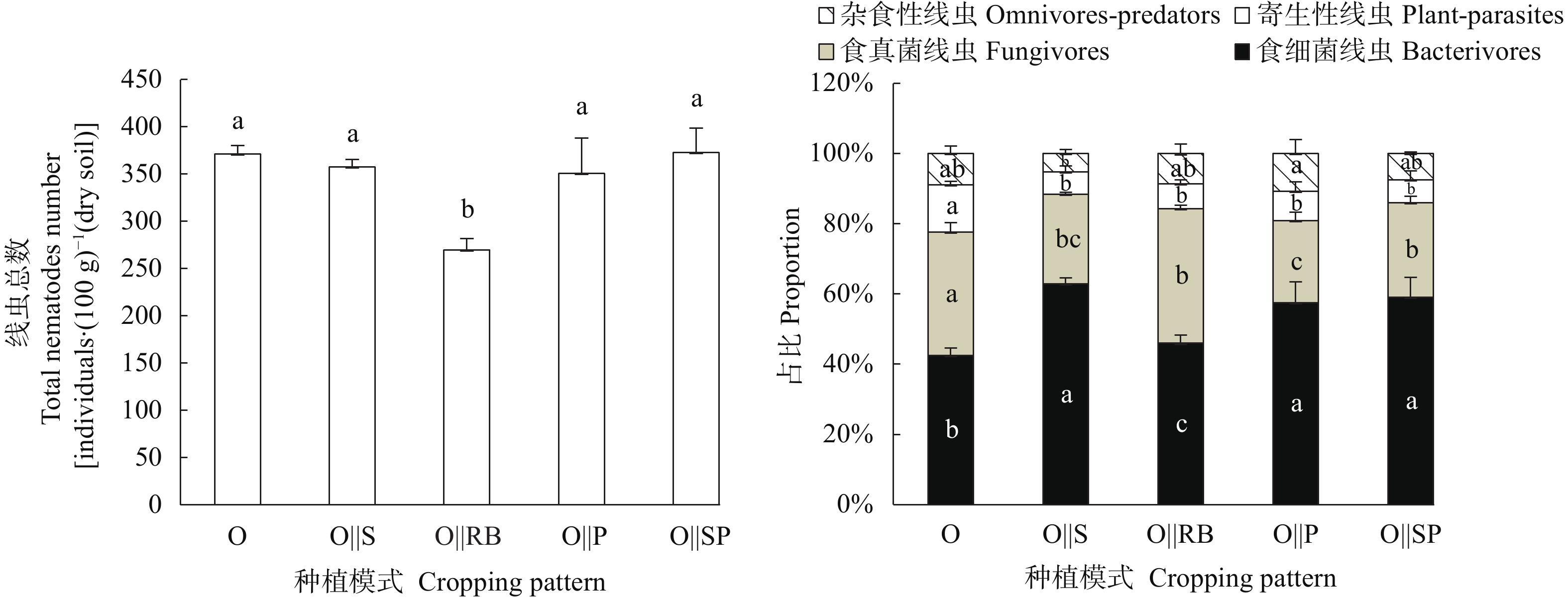
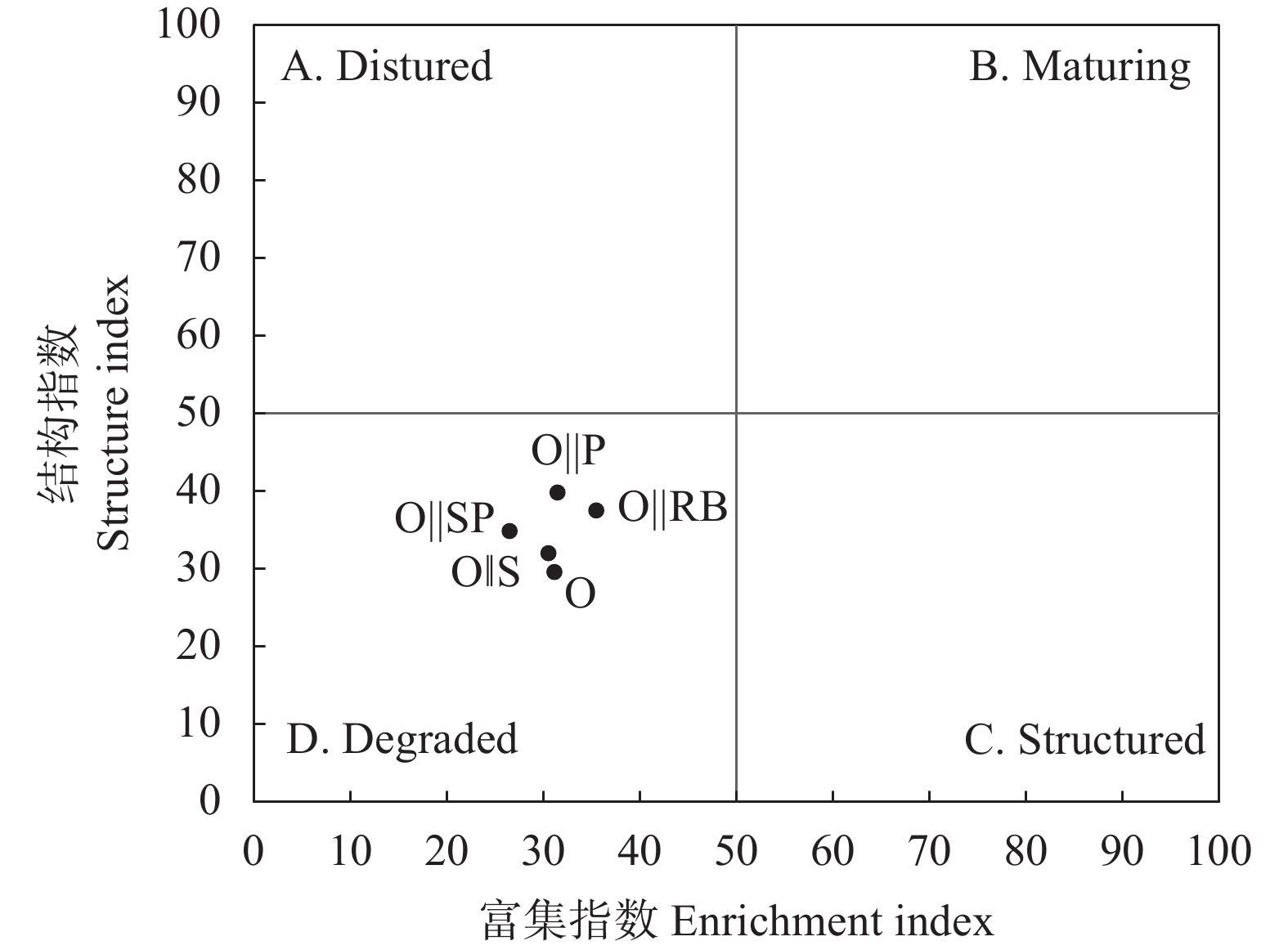
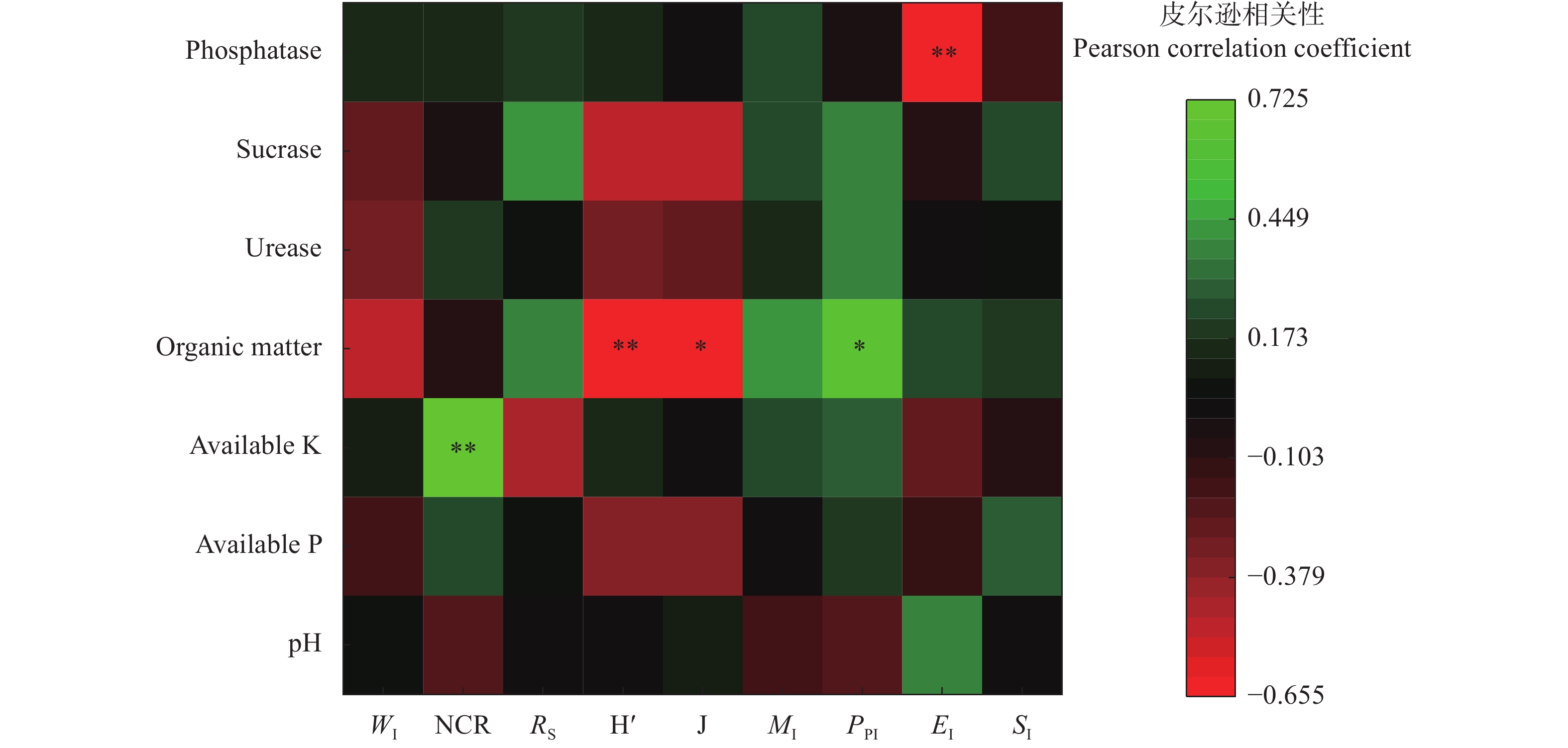
间作是土地可持续利用的重要种植模式。土壤线虫作为土壤健康的指示生物, 可揭示地下生态系统的食物网功能。为探究燕麦不同间作模式对产量和土壤线虫群落的影响, 本试验分别设置燕麦单作、燕麦||大豆、燕麦||赤小豆、燕麦||马铃薯、燕麦||甘薯5个处理, 分析燕麦不同间作模式对作物产量以及土壤线虫的数量、多样性和群落结构的影响, 以筛选出较优的燕麦间作模式。结果表明: 相比单作, 间作模式在土地利用率和作物总产量方面有一定的优势, 其中燕麦||大豆间作模式最佳, 土地当量比(1.56)最高。4种燕麦间作模式均显著(P<0.05)降低了植物寄生线虫的相对丰度, 提高了食微线虫相对丰度, 优化了土壤线虫群落结构。燕麦||大豆间作模式下食微线虫比例最高(88.42%), 植物寄生线虫比例最低(6.31%), 且在多项生态指数中(瓦斯乐斯卡指数、线虫通路指数、多样性指数和均匀度指数)显著高于燕麦单作(P<0.05)。而燕麦||赤小豆间作模式下富集指数最高, 植食性线虫成熟度指数最低, 土壤线虫群落受干扰程度最低。速效钾与线虫通路指数呈极显著正相关(r=0.722**), 可以使土壤有机质分解主要依靠细菌分解途径。综上, 间作种植模式可以优化土壤线虫群落结构, 改善土壤生态环境, 提高产量; 燕麦||大豆是该试验条件下最佳的间作模式。
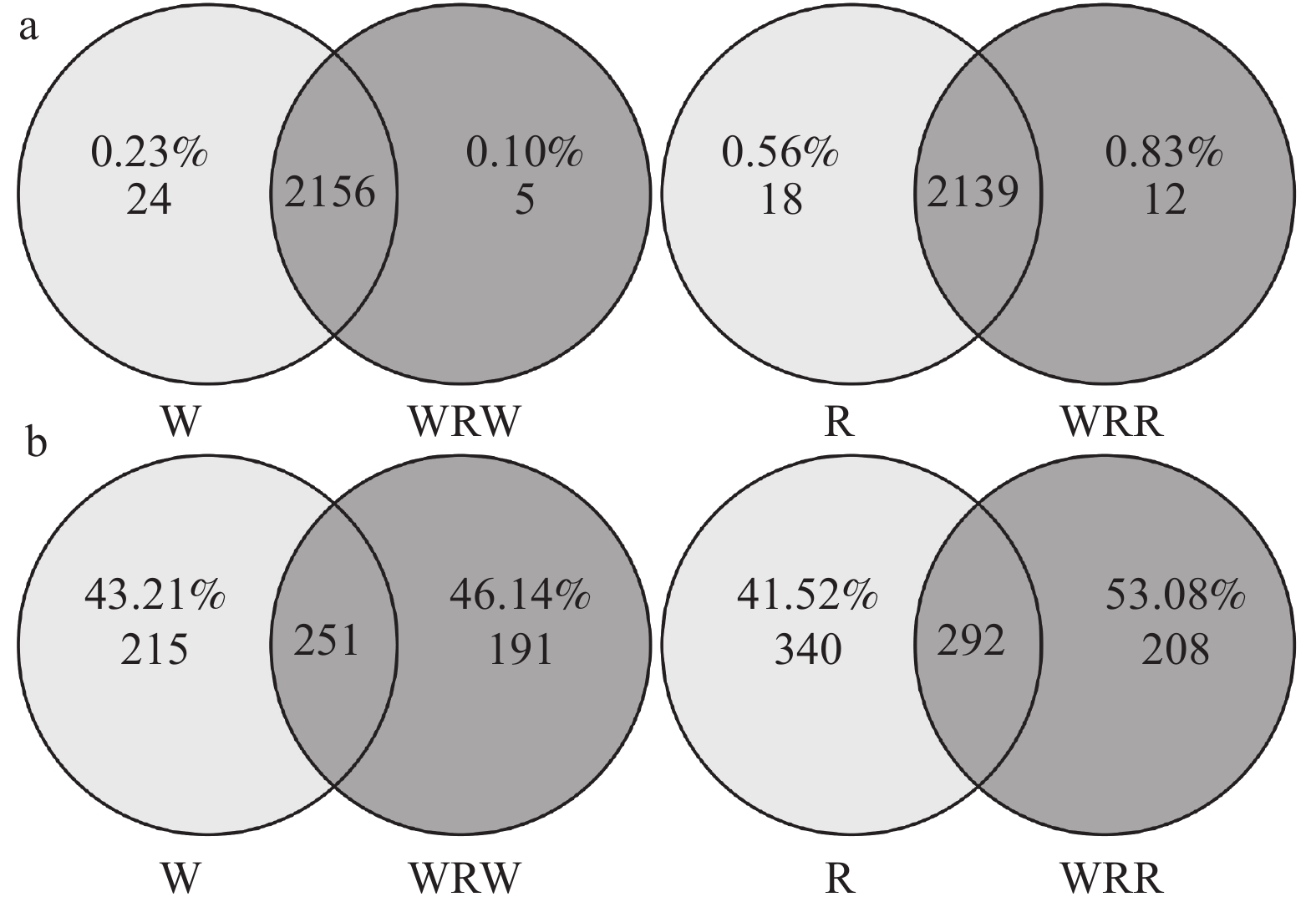
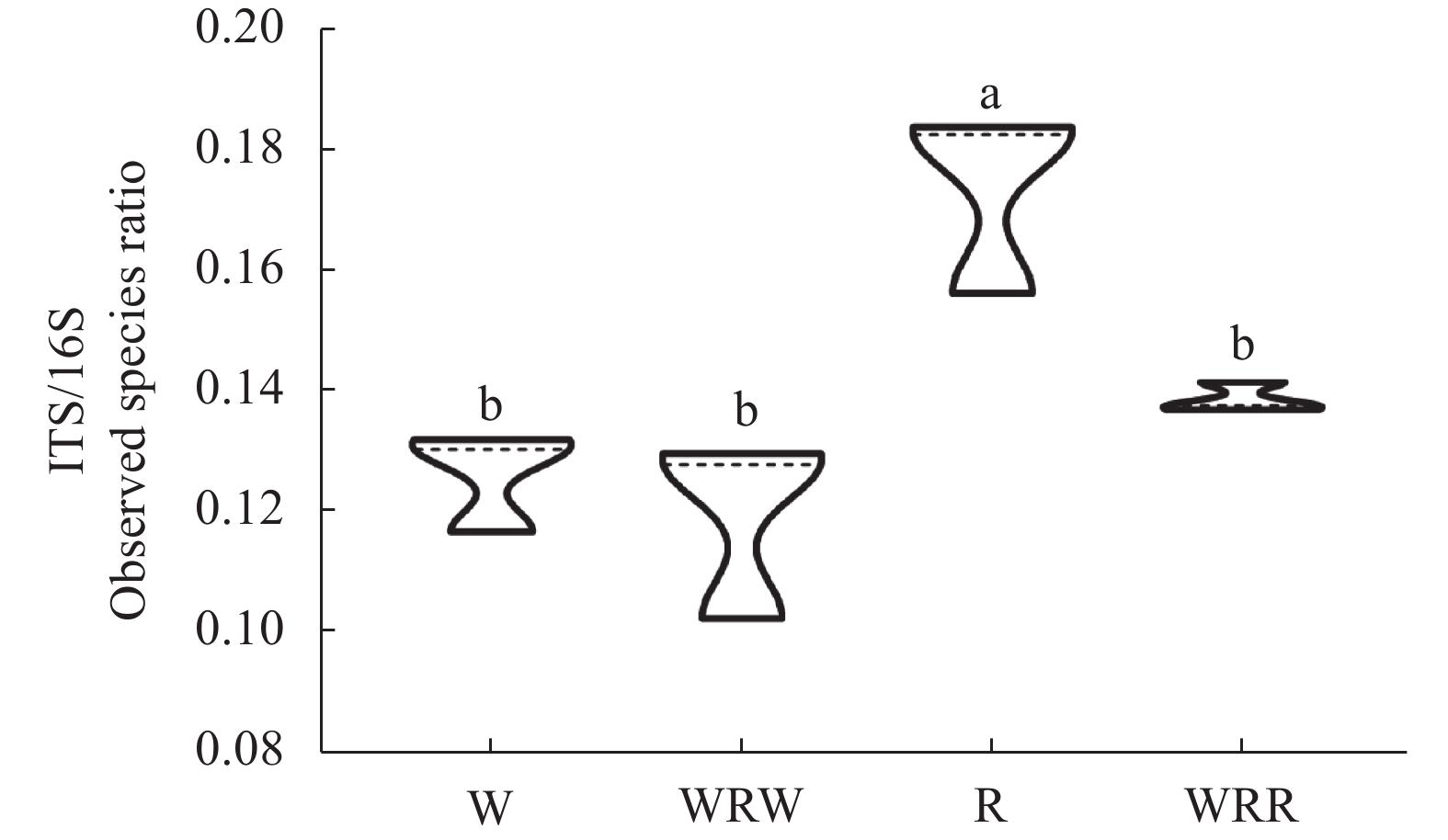
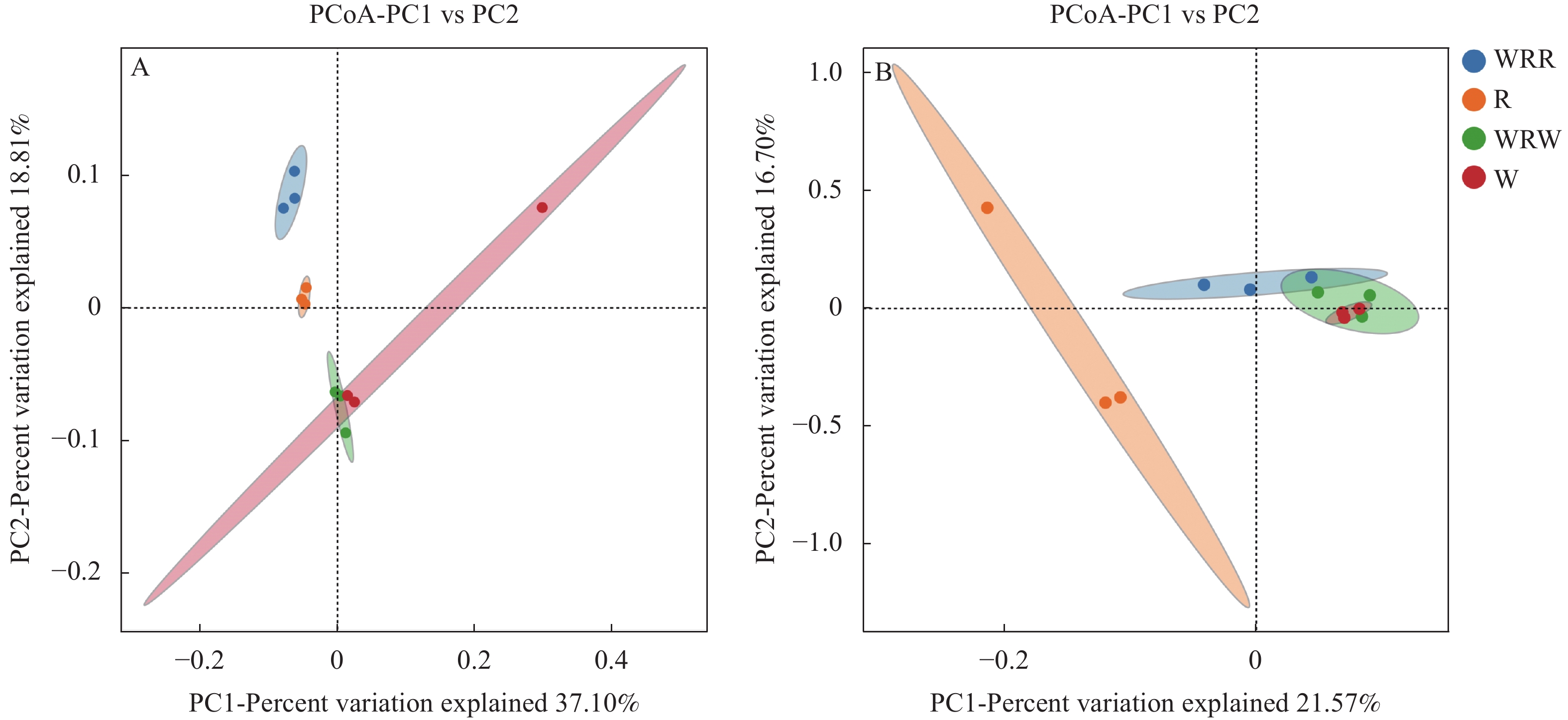
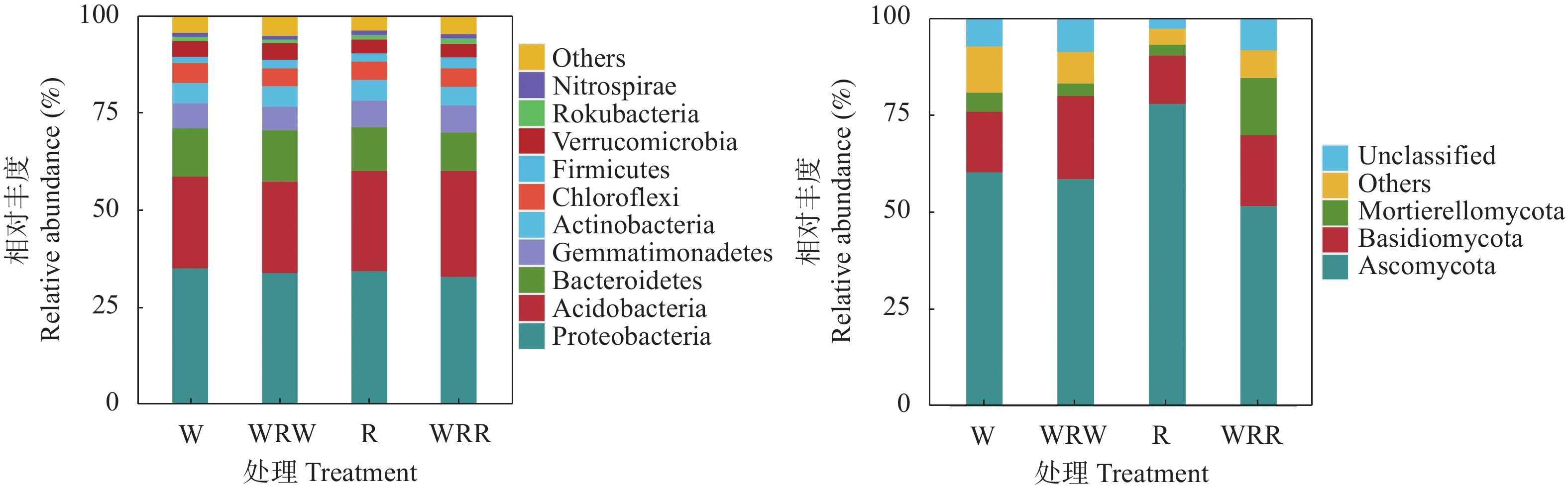
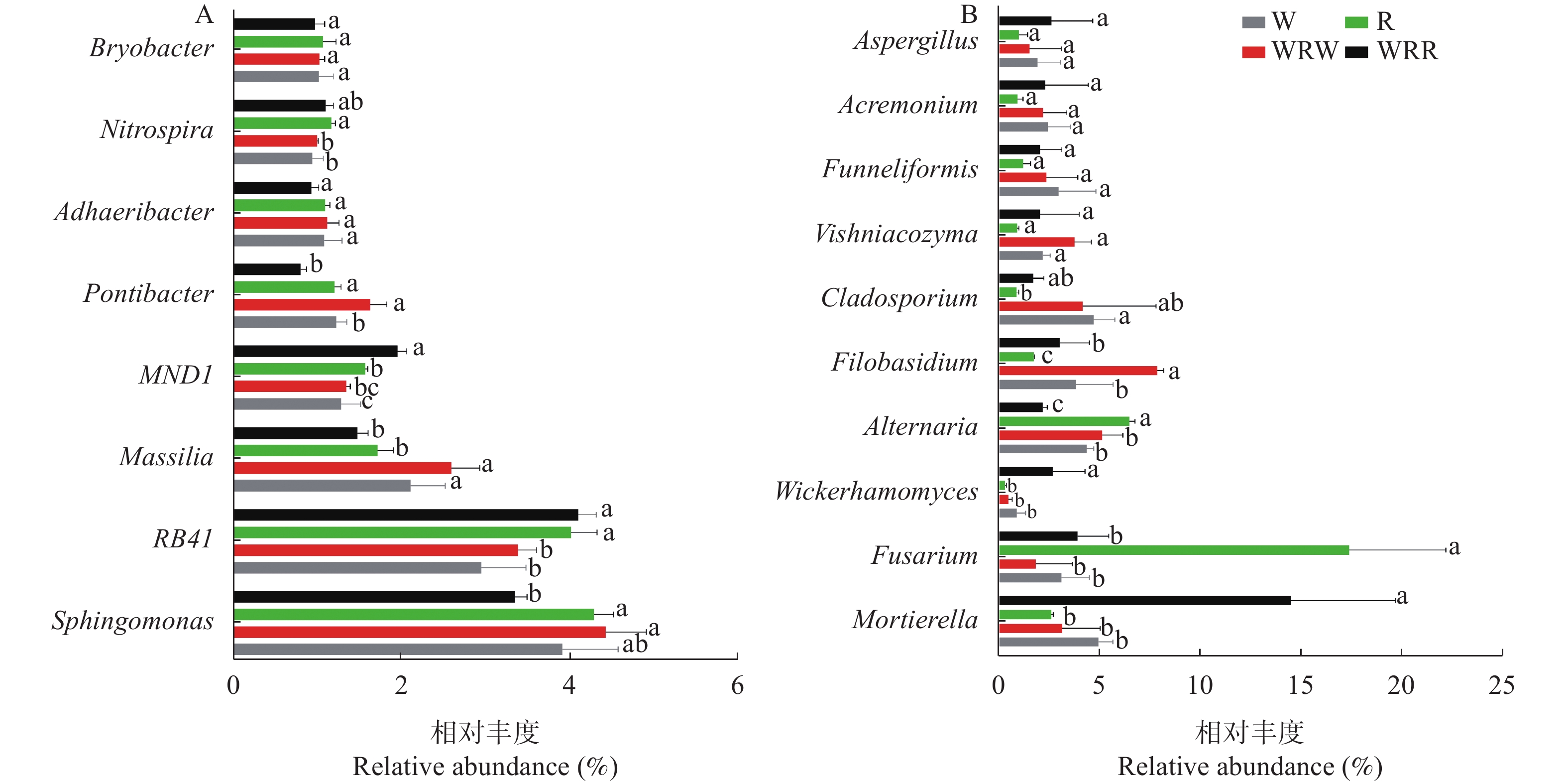
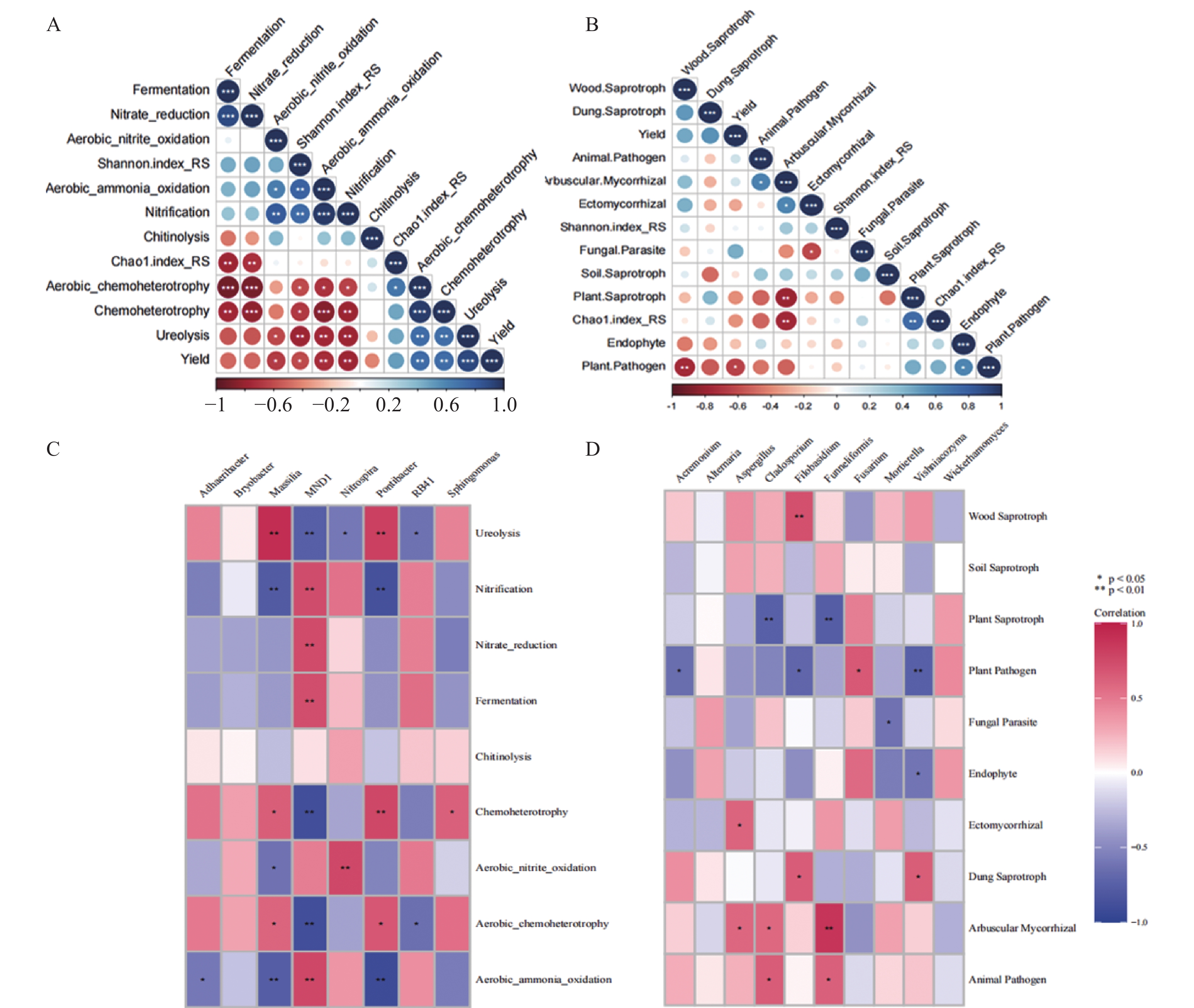
研究小麦||红花间作系统作物根际土壤微生物群落结构及功能变化, 对后期构建合理的小麦||红花种植模式和地区适宜性评价具有重要意义。本研究在田间设置小麦红花间作(2∶1)模式, 以小麦单作、红花单作为对照, 基于Illumina Miseq高通量测序平台研究了小麦||红花根际土壤细菌和真菌群落结构特征和功能变化, 在成熟期, 分别测定小麦红花产量。结果表明: 小麦||红花土地当量比为1.01, 间作优势并不明显; 小麦、红花根际微生物多样性指数分析结果表明, 间作提高了小麦根际细菌多样性和真菌的丰富度, 但显著降低了小麦根际细菌的丰富度和真菌多样性;间作显著提高了红花根际细菌的多样性, 但降低了红花根际真菌的多样性及细菌和真菌的丰富度。主坐标分析(PCoA)和群落均衡性分析结果表明, 间作显著改变了红花根际微生物群落结构(P<0.05)。小麦红花根际优势细菌门为变形菌门、酸杆菌门和拟杆菌门, 在单作小麦、间作小麦、单作红花和间作红花中平均相对丰度总占比分别为65.94%、70.57%、71.39%、70.07%, 优势真菌为子囊菌门、担子菌门, 平均相对丰度总占比分别为75.99%、68.17%、93.23%、69.88%。间作显著提高了红花根际酸杆菌门、Rokubacteria、以及真菌的被孢霉门的相对丰度(P<0.05), 对小麦各菌门并未造成显著影响。功能预测结果表明, 间作显著提高了红花根际好养氨氧化、硝化作用、发酵、硝酸盐还原等养分循环过程(P<0.05), 结合相关性分析结果发现MND1与这些过程显著正相关(P<0.05), 且其相对丰度在红花根际表现为间作显著高于单作(P<0.05)。此外, 间作显著提高了红花根际丛枝菌根功能基因的相对丰度, 同时降低了植物病原菌类功能基因的相对丰度(P<0.05); 在属水平, 间作显著提高了红花根际被孢霉属、Wickerhamomyces等有益菌属的相对丰度, 显著降低了镰刀菌属、链格孢属致病真菌属的相对丰度(P<0.05)。共线网络分析结果表明, 间作可一定程度上提高小麦根际微生物网络的复杂性, 降低红花根际微生物网络的复杂性;此外, 镰刀菌属位于红花根际微生物网络的中心, 间作可通过影响红花根际微生物间的相互作用明显降低镰刀菌属的数量。综上, 小麦||红花间作体系中, 处于竞争劣势的红花可通过改善根际微生物群落组成, 提高根际养分循环过程, 进而缓解在养分等资源竞争上的不利地位; 此外, 间作可显著降低红花根际致病类病原菌的相对丰度, 对减少红花病害的发生及土壤中致病菌总量均具有重要意义。
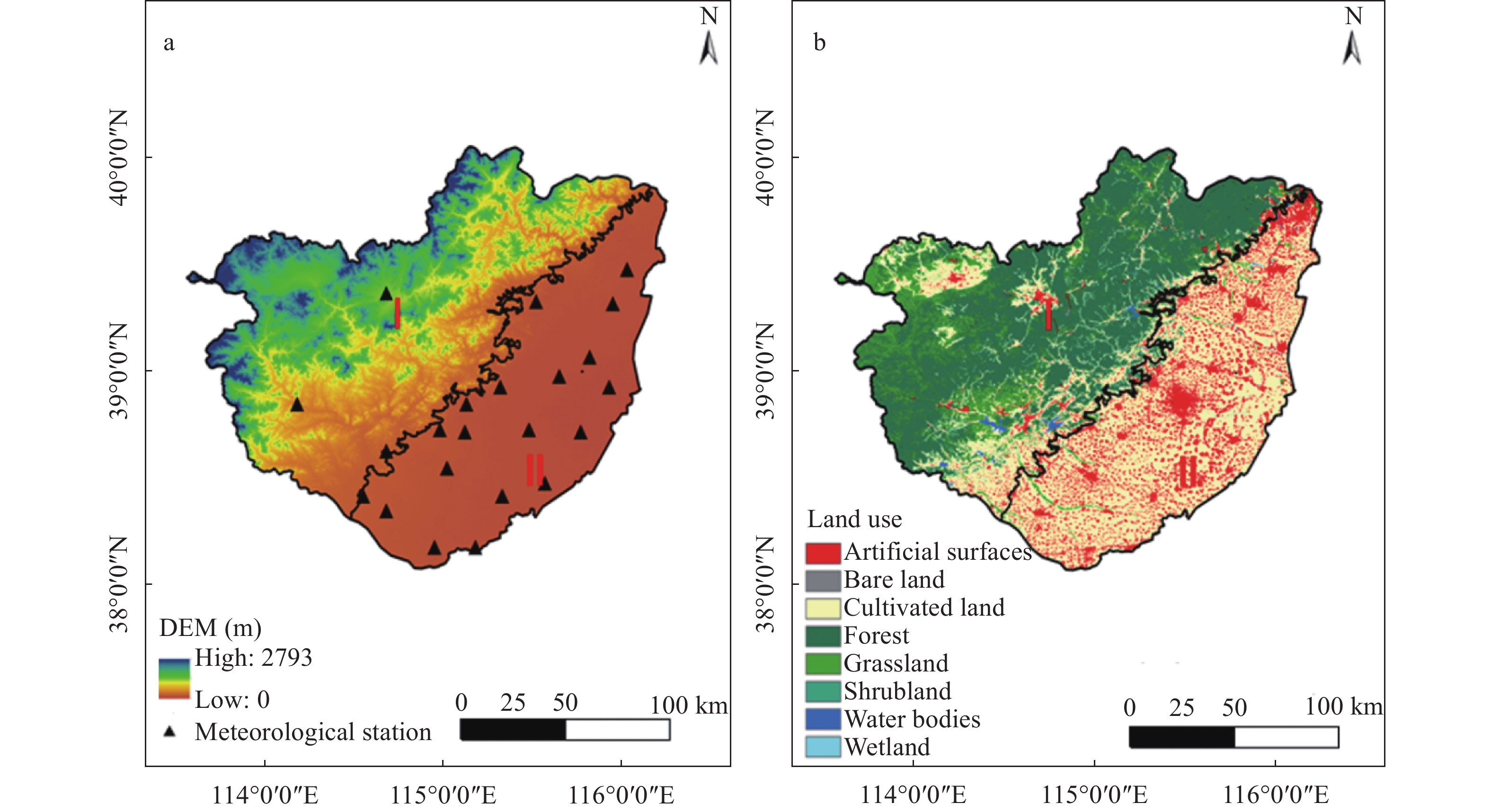
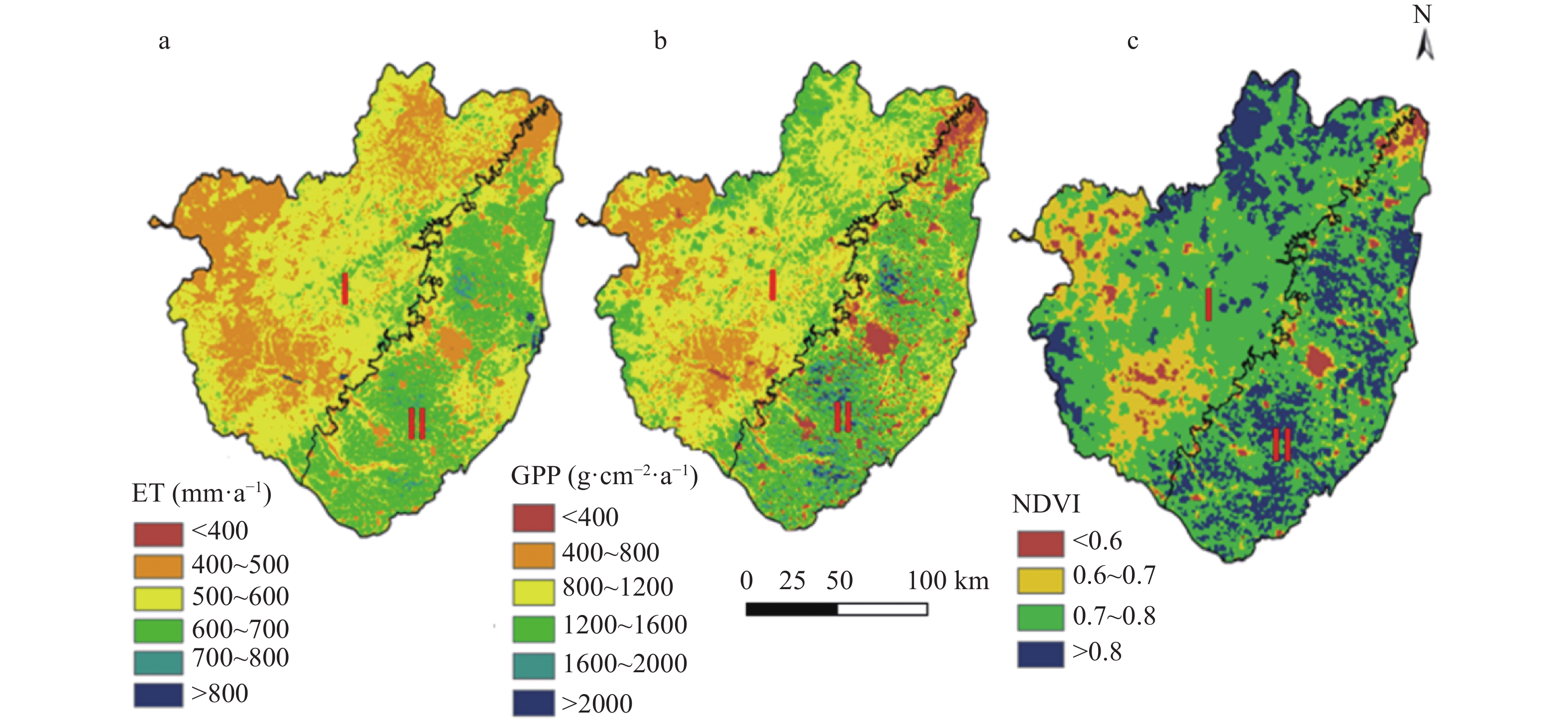
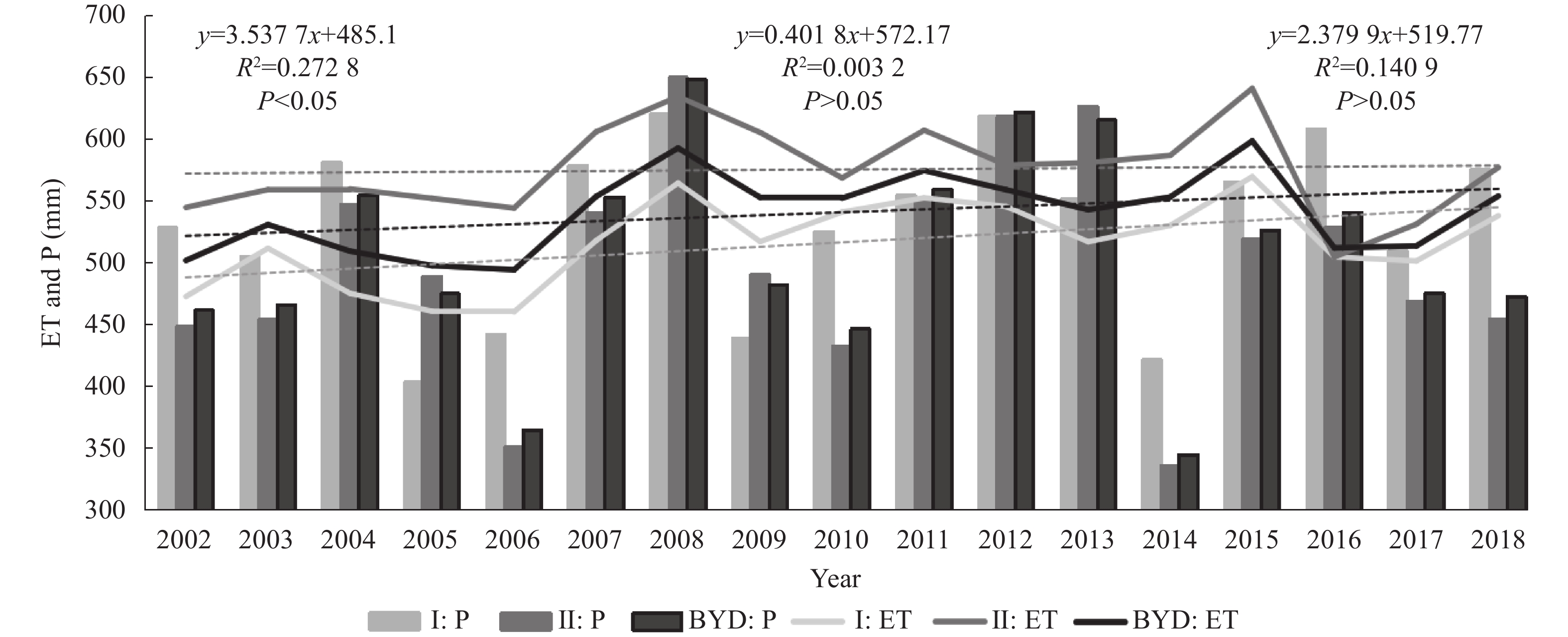
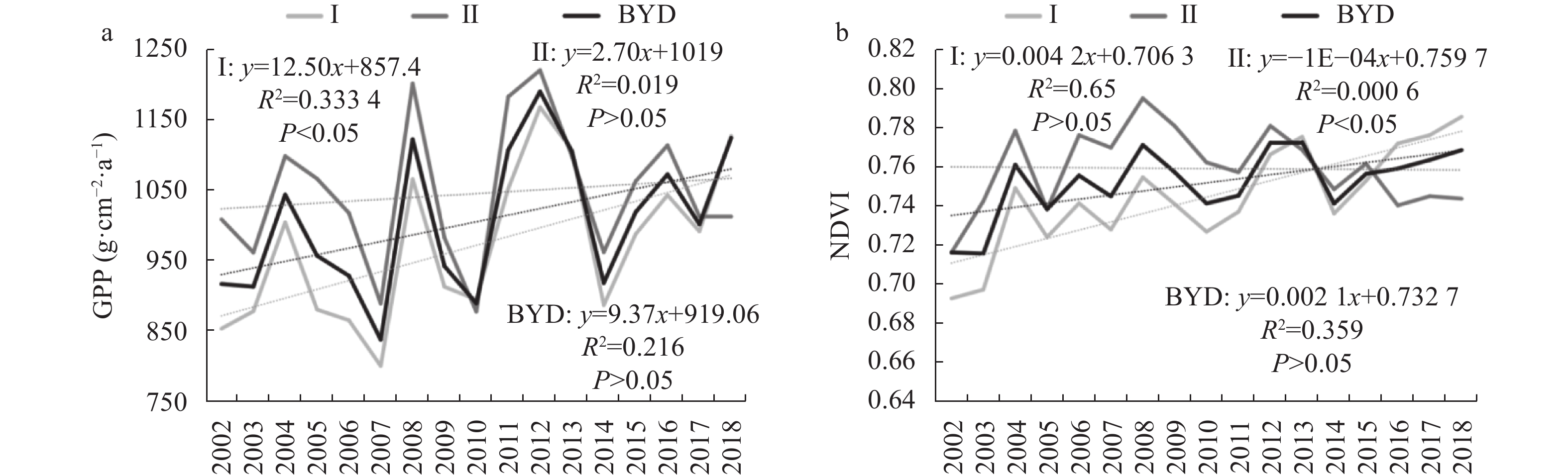
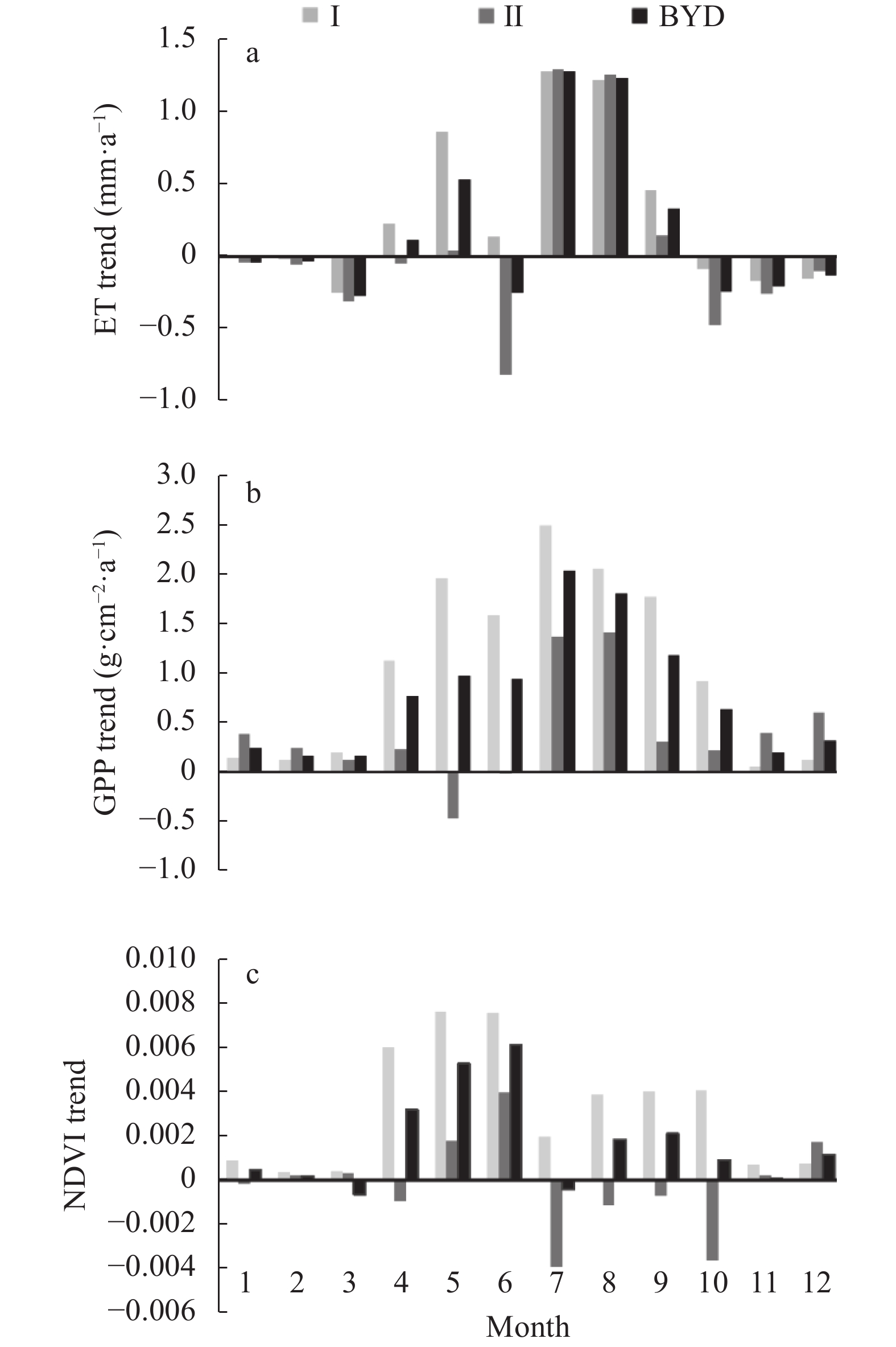
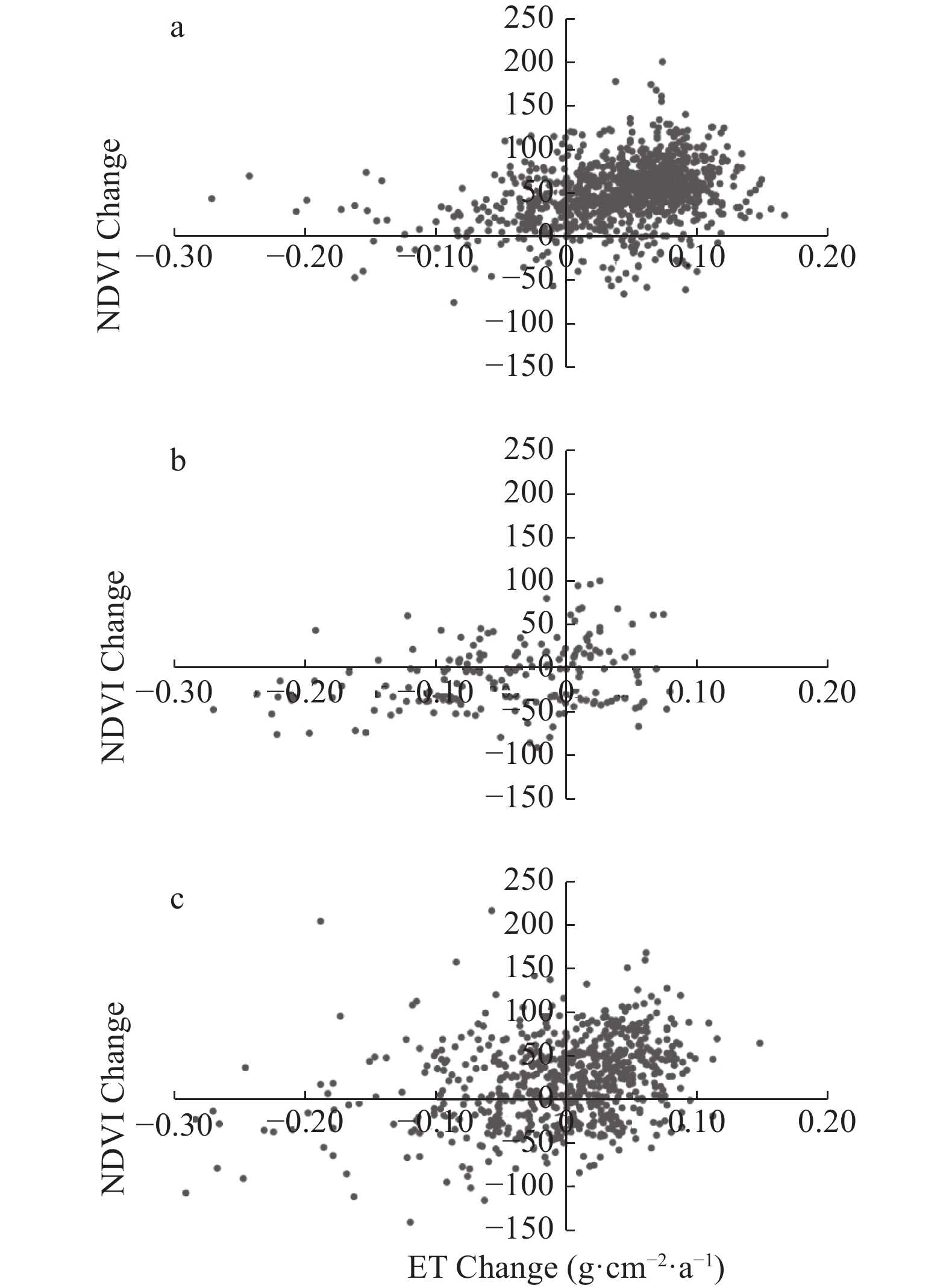
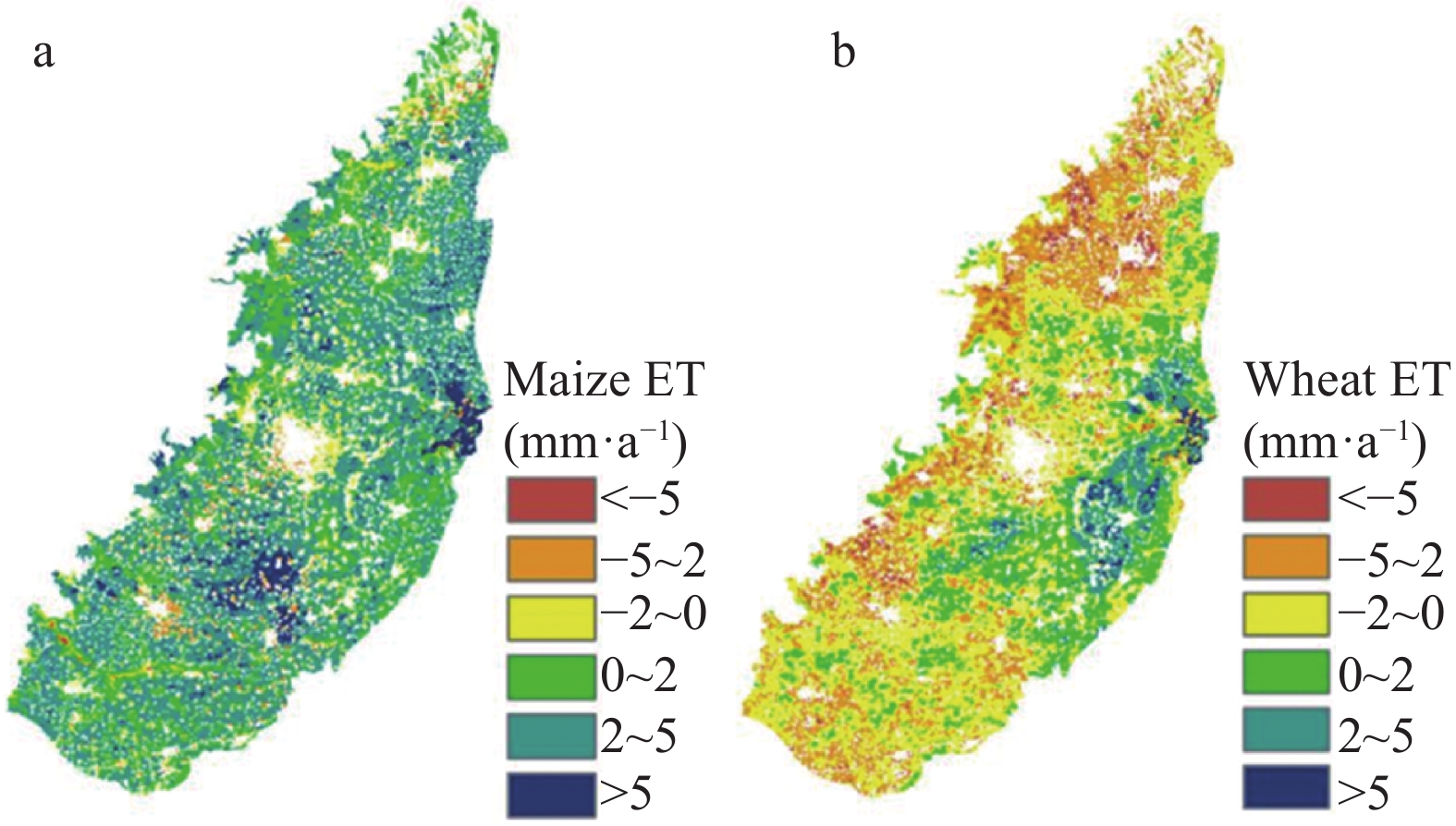
Baiyangdian Catchment is facing a growing shortage of water resources. Identifying the sensitive drivers of evapotranspiration (ET) changes from land and crop management will be critical to understanding the reasons for mountainous runoff reduction and depletion of groundwater resources in the plain. It will also be important for making Xiong’an become a Future Example City for green and sustainable development. In this study, remotely sensed ET data from PML V2 products with a spatial resolution of 500 m was used to analyze the trend of ET at the pixel level and to understand its influence on vegetation such as GPP (Gross Primary Production) and NDVI (Normalized Difference Vegetation Index) under different land-use types for 2002‒2018. Results showed that there was a significant increase in ET in mountain regions and a slight increase in plain regions of the catchment. The spatial pattern of mean annual ET was very much relevant to the changing trend of GPP and NDVI. For the whole catchment, the average increases of ET, GPP, and NDVI were respectively 2.4 mm, 9.8 g∙cm−2, and 0.0021 at an annual rate. In the mountainous region, changes in annual precipitation and vegetation recovery together caused a total increase of ET by 56.5 mm over the period and negatively affected the runoff. In the plain region, there were 3 factors influencing the change of ET. While intensification of urbanization and reduction in the cultivation of wheat, the water consumptive crop, had both resulted in the decrease of ET and water consumption, ET or water consumption in most irrigated fields increased. Since the beneficial effects from urbanization and crop adjustment were not enough to offset the increase of ET in irrigated fields, an overall ET increase of 6.4 mm over the period was found. In conclusion, both in the mountainous and plain regions, ET increased. And therefore, more efforts are needed to control the ET increase in natural vegetation and cropland for a green and sustainable catchment.

碳排放精准测算是“双碳”目标实现的重要保障。在进行农业碳排放测算时, 作物类型、生产方式、地理区位等因素均会对碳排放产生影响, 因此, 虽然诸多学者尝试从不同角度对中国农业碳排放展开测度, 但在估计方式、样本选取和测算结果等方面仍未形成一致的有效结论。本研究首先介绍了中国农业碳排放的主要核算方法, 包括排放系数法、模型模拟法和实地测量法; 其次, 从投入产出、生产过程、碳汇以及碳足迹4个方面对现有农业碳排放核算方式进行梳理; 再次, 对农业碳排放的核算结果进行归纳总结; 最后, 分析现有研究的局限性, 并对未来农业碳排放核算研究进行了展望。研究发现: 现有研究在农业碳排放测算时, 存在遗漏排放源、排放系数使用不科学以及研究视角过多集中于宏观层面等不足。建议未来从构建科学全面的农业碳排放核算体系、完善排放系数以及加强农户等微观层次研究等方面展开。
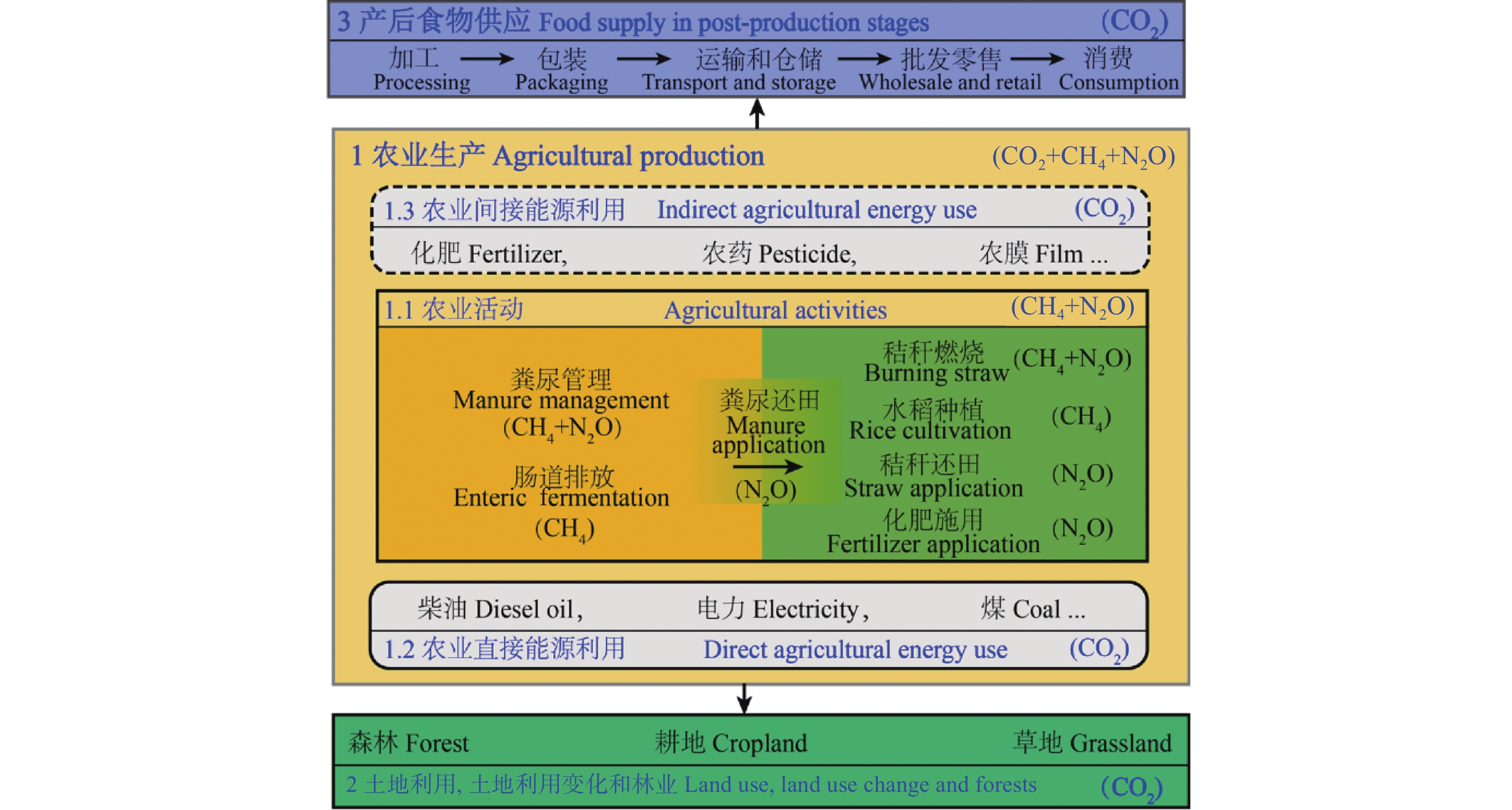
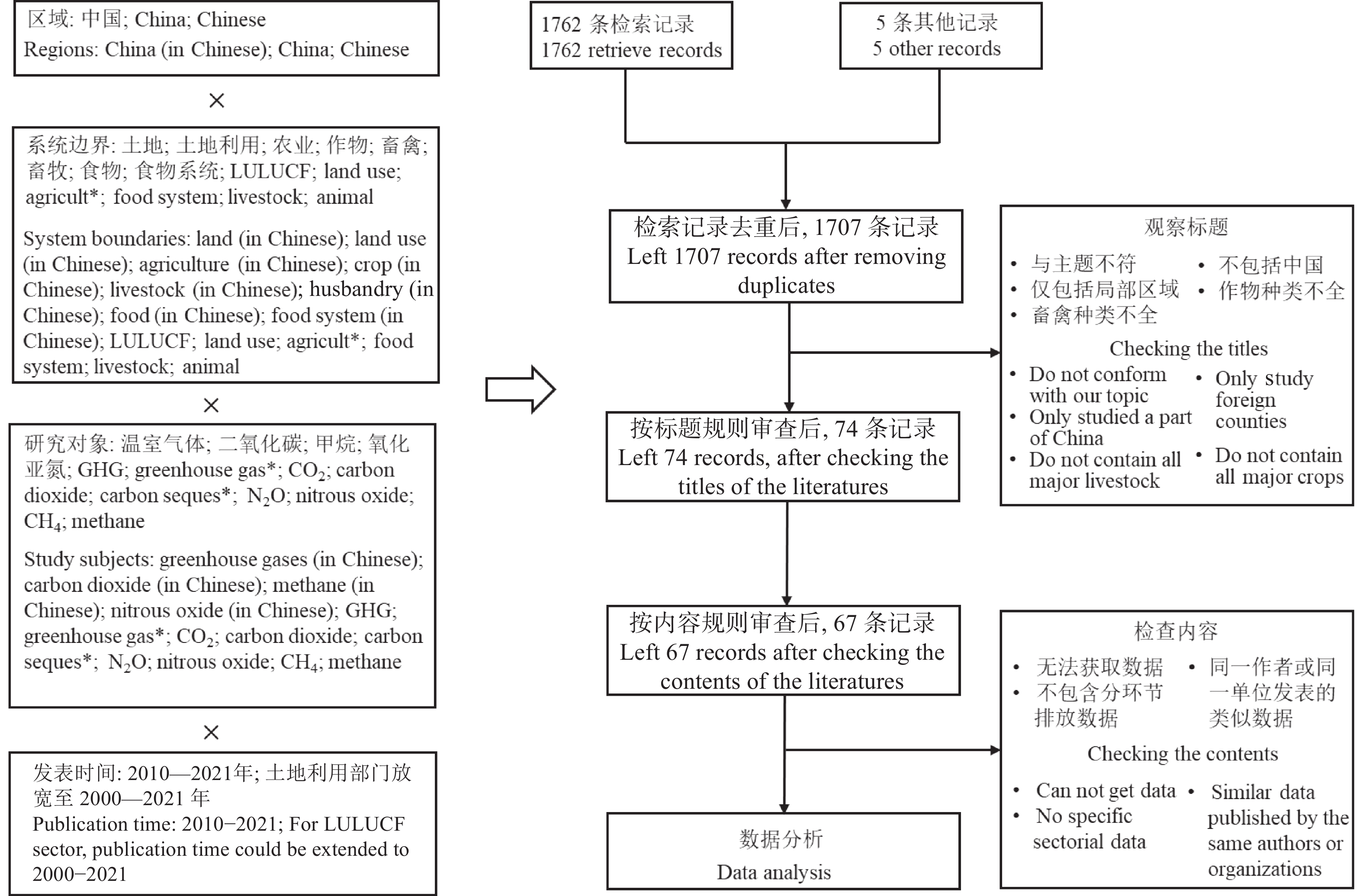
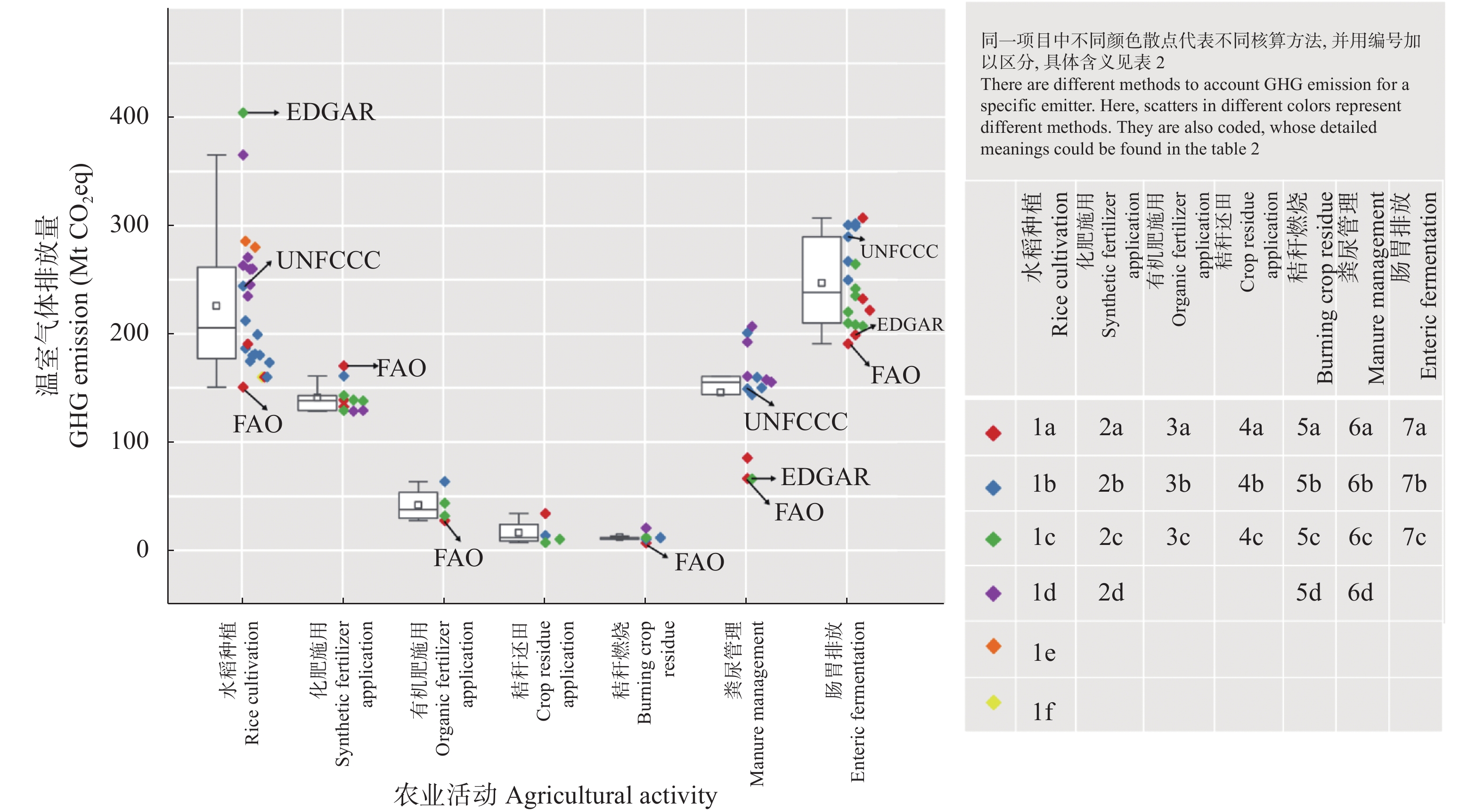
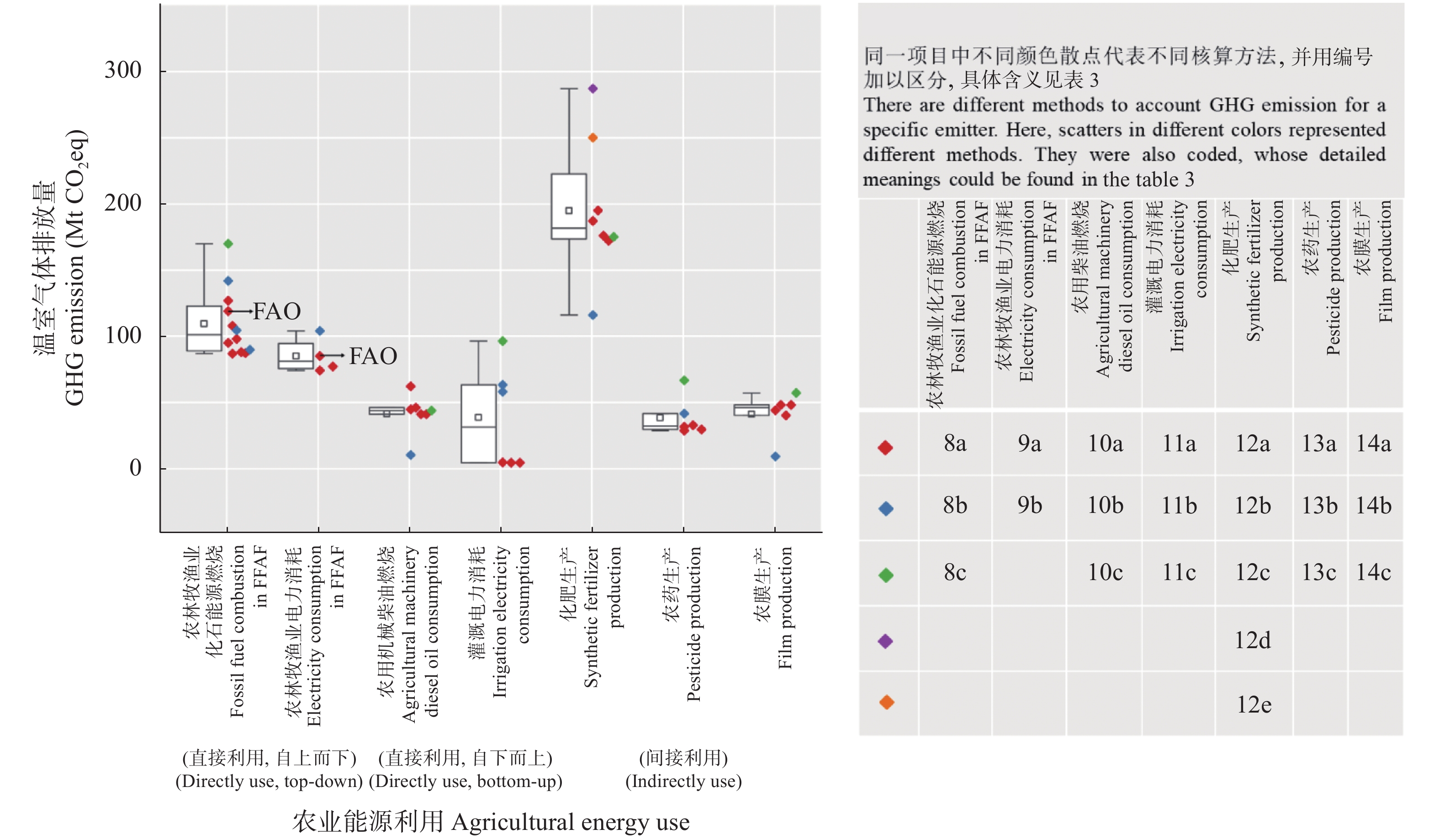
我国食物系统温室气体核算研究相对缺乏, 相关研究多散见于农业和畜牧业部门, 难以满足碳达峰、碳中和背景下减排和固碳的需求。本文提出了一个涵盖土地利用、土地利用变化和林业(LULUCF)、农业生产、产后食物供应部门的食物系统温室气体排放与吸收的核算框架; 通过文献综述、温室气体排放量收集和排放参数推算, 剖析了各环节排放核算方法的差异和排放参数的不确定性。结果表明, 粪尿和秸秆还田、农药和农膜生产、食物加工、批发和零售以及草地的碳排放(或吸收)参数的变异系数(CVs)在35%以上。未来食物系统温室气体核算研究需: 1)在农业生产部门, 细化农业活动排放参数, 整合基于终端能源消费和农业生产过程的温室气体核算方法, 并加强农业投入品能耗研究; 2)在LULUCF部门, 建立适用于全球变化研究的土地利用分类体系, 识别食物系统相关的土地利用过程, 将实地调查核算法和基于过程模型的核算方法相互校验; 3)在产后食物供应部门, 明确各环节排放的核算范围, 有针对性地选择投入产出-生命周期评价法(EIO-LCA)、基于过程的生命周期评价法(PLCA)与终端能源消费核算法等。本文可进一步为食物系统温室气体减排提供科学依据。
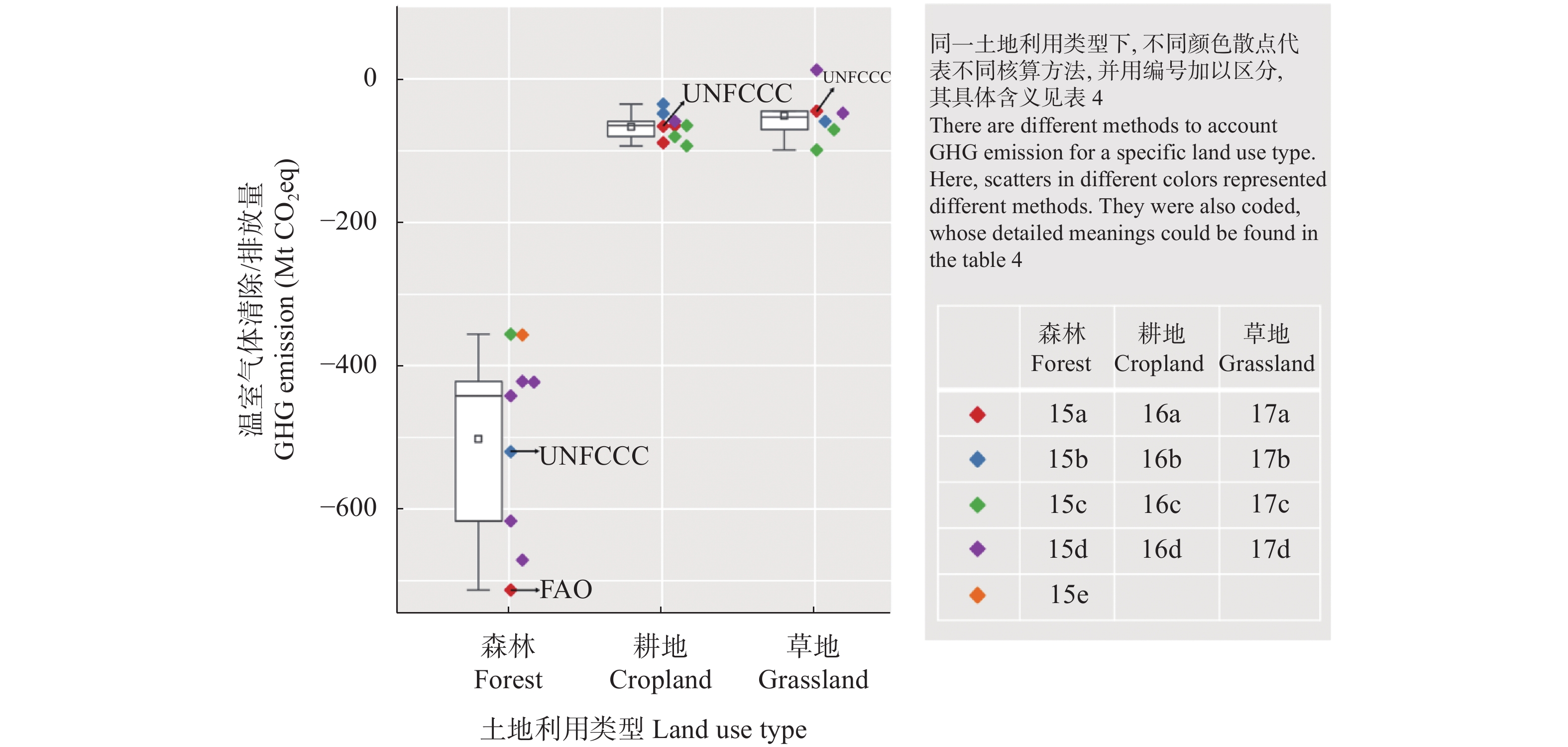
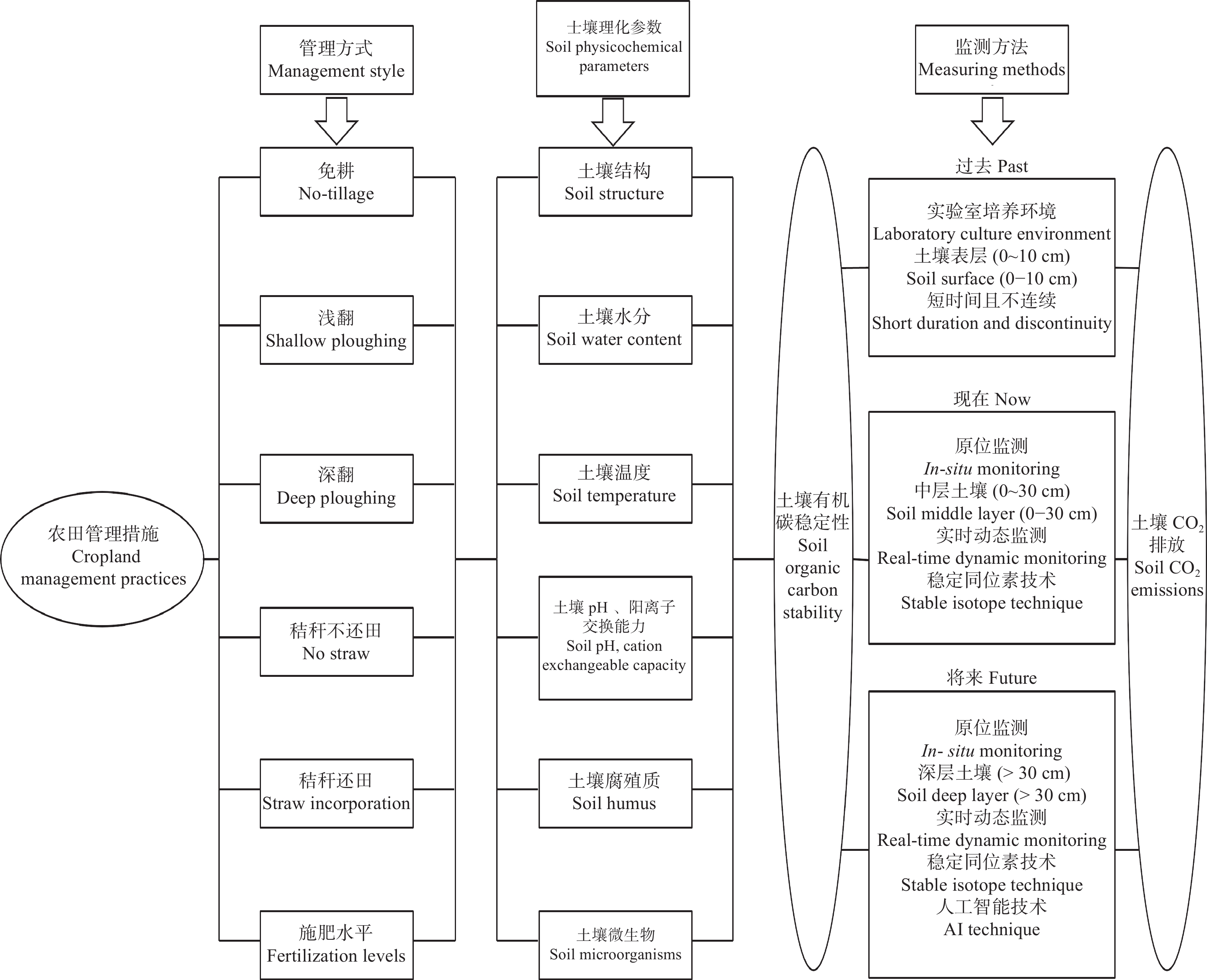
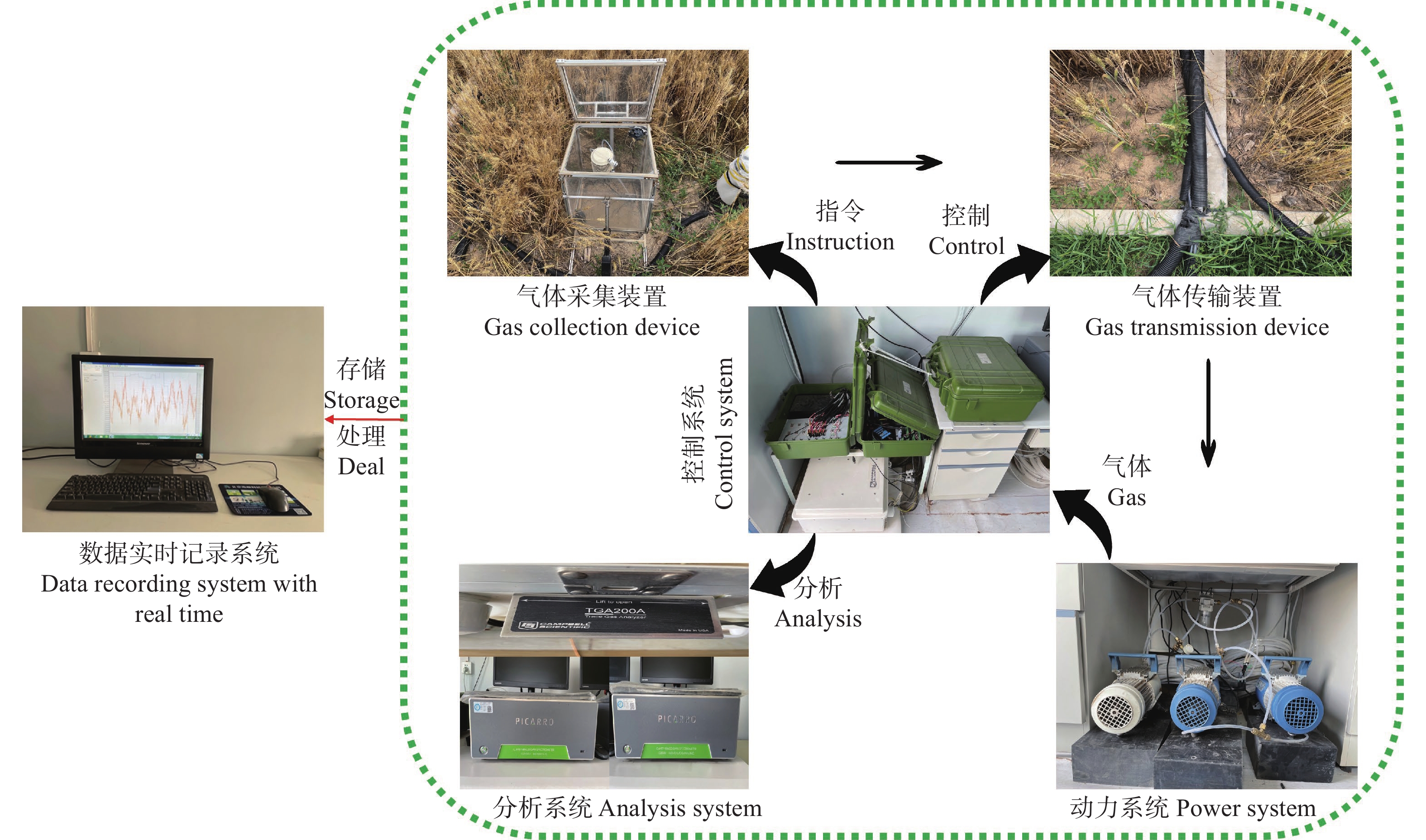
土壤有机碳是地球表层储量最高且储存周期最长的生态系统碳库之一。如何提高土壤有机碳稳定性和增强土壤固碳减排能力, 是陆地生态系统碳管理可持续战略的关键科学问题。国际学术界一致认为农田生态系统在固碳方面的作用越来越明显, 在实现碳中和的进程中发挥重要的作用。农田管理实践方式会扰动土壤碳循环过程, 采取有效的管理方式会使其成为碳汇。目前, 国内研究主要集中在耕作方式、施肥和灌溉水平、秸秆还田对农田生产力、碳固持速率、温室气体排放方面的影响, 但就农田生态系统有机碳稳定性对不同农田管理方式的响应机制以及与土壤碳排放之间的关系认识尚未明确。13C同位素技术是研究农田土壤碳循环过程的有力工具, 通过测定土壤碳排放过程中不同有机碳组分的同位素丰度, 能够精准区别土壤呼吸组分和来源, 从而更好地揭示土壤有机碳稳定性对农田管理措施的响应机制, 为增强土壤碳汇效应和农业可持续发展提供科学依据。以往的研究大多集中在模拟试验以及小范围、短时间监测, 与实际差距较大, 测量结果会高估或低估实际值。因此, 在未来的农田土壤碳循环研究过程中要采取多点、长时间实时原位监测, 并结合13C同位素技术, 实现土壤CO2排放实时分解, 达到揭示土壤有机碳稳定性机制的目的。

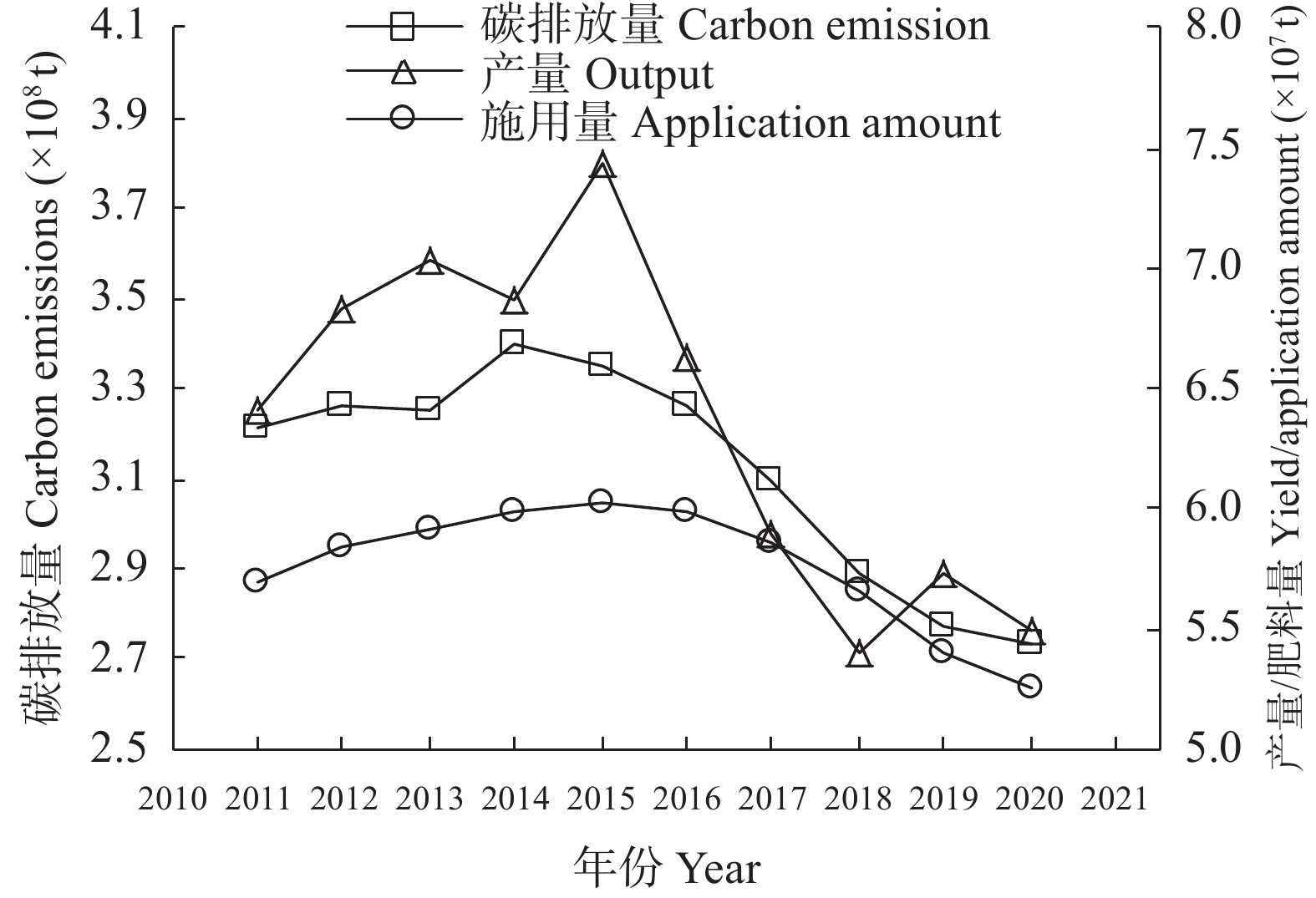
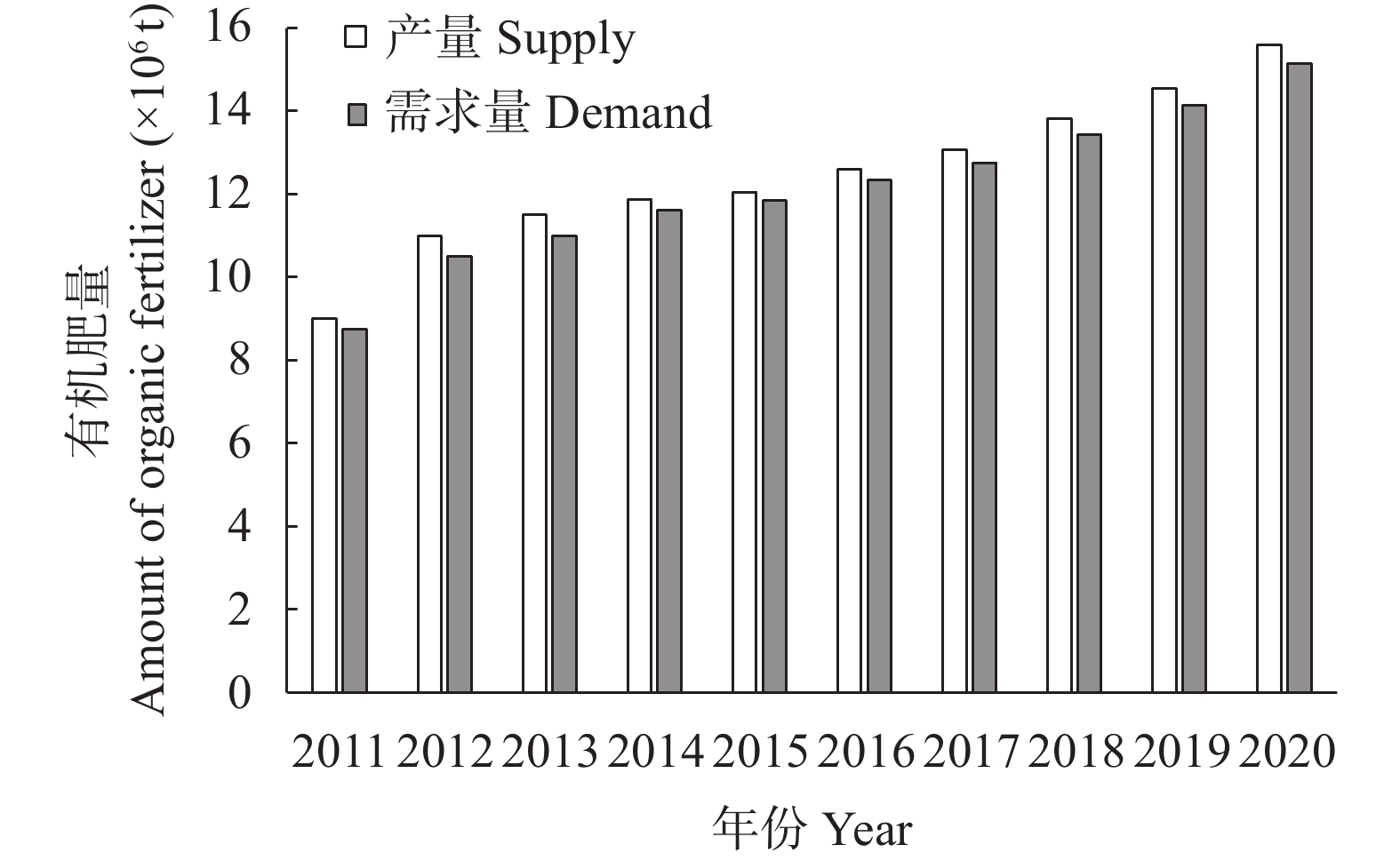
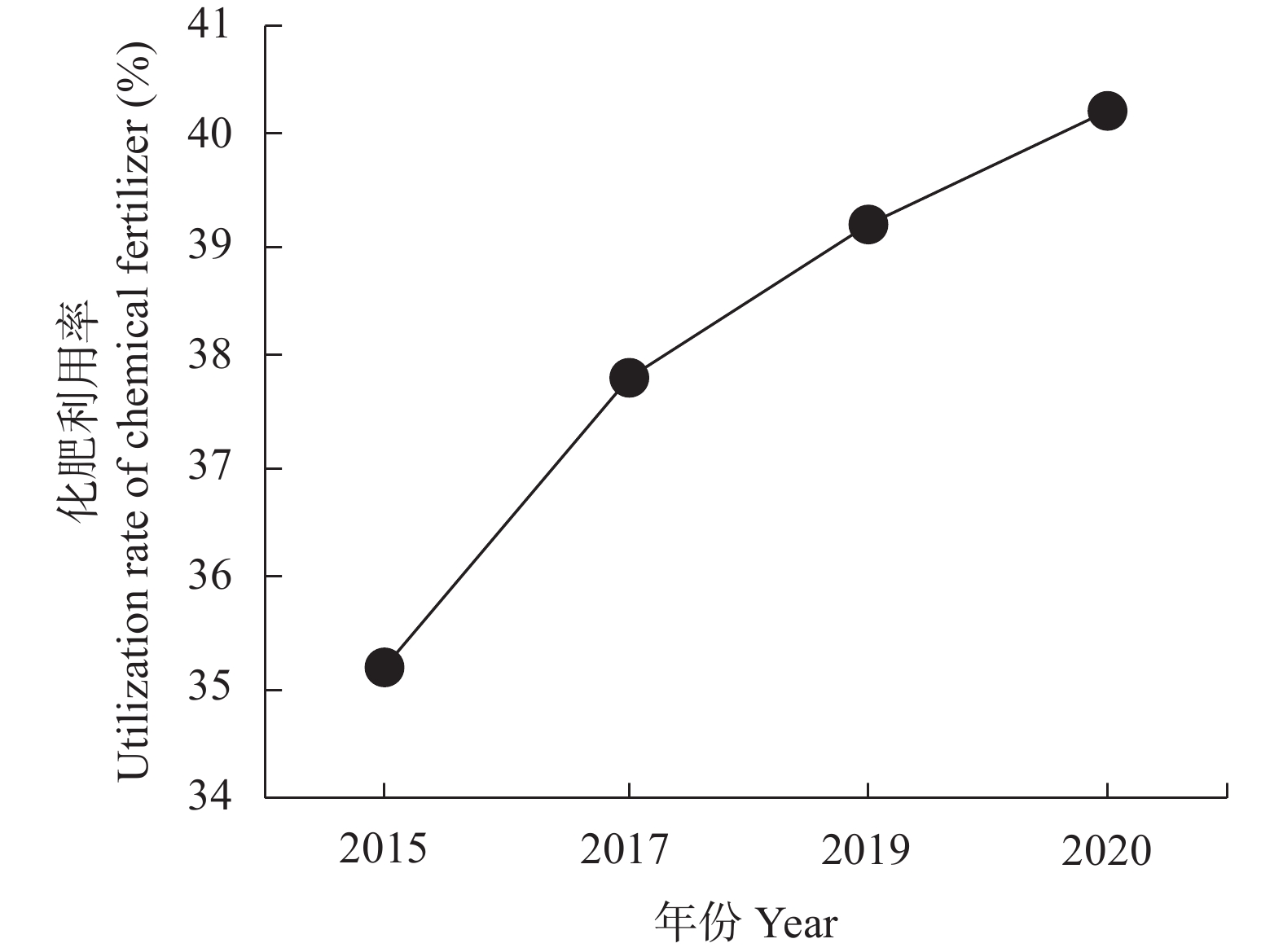
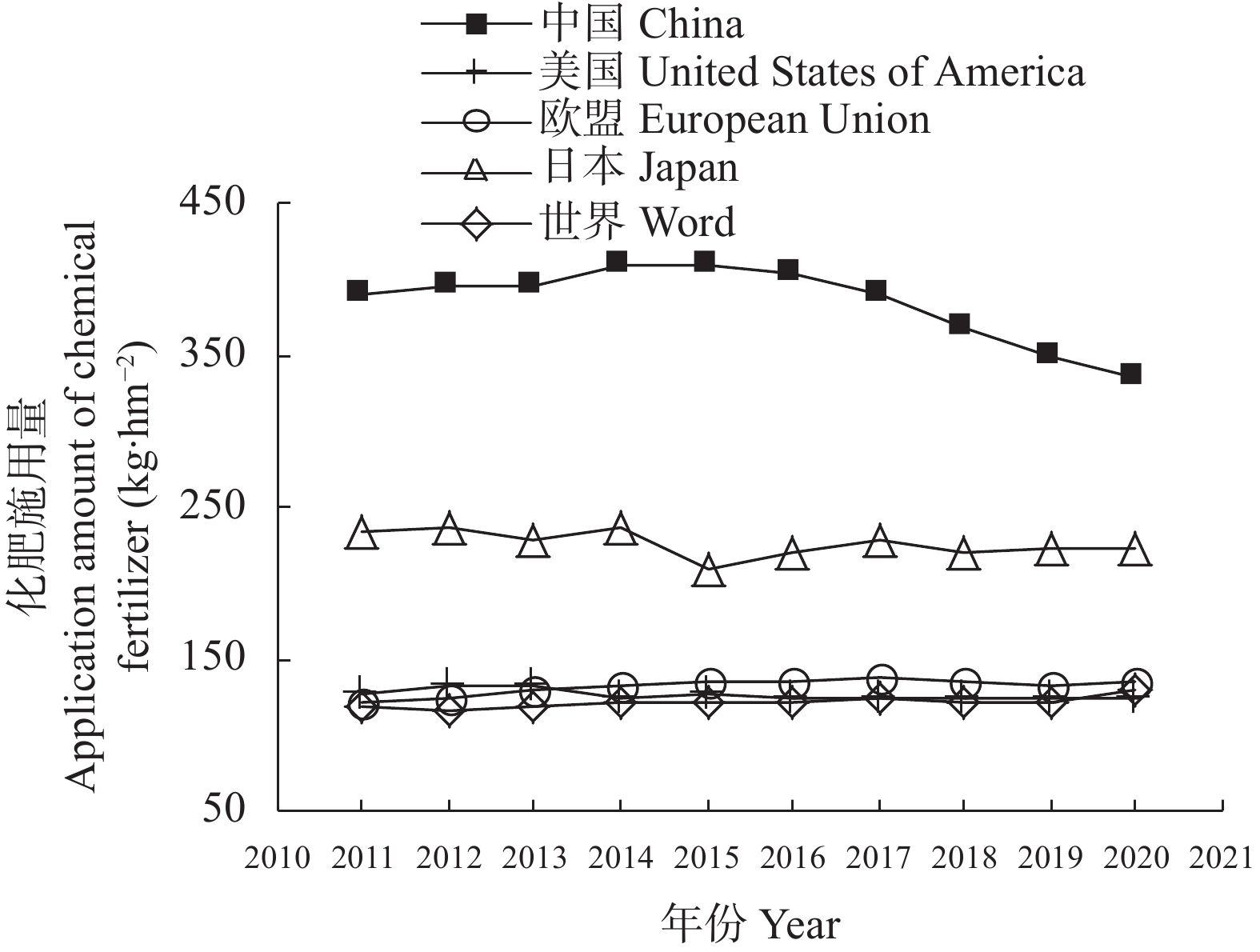
碳达峰和碳中和目标已经成为世界发展的共同目标。肥料在碳排放中起着双重作用, 既消耗能量产生碳排放, 又提升作物固碳能力减少碳排放。未来很长一段时间, 肥料对于粮食和农业生产仍具有不可替代作用, 使用合适的肥料产品、科学施用肥料, 对于减少碳排放有重要的作用。本研究利用联合国粮农组织、国家统计局、农业农村部等网站公布的数据资料, 分析我国肥料行业碳减排现状, 提出存在的问题, 探讨“双碳”目标下肥料行业的发展建议, 以期为肥料行业低碳化发展提供参考。结果表明: 2011—2020年我国肥料行业碳减排成效显著。化肥产量和施用量呈先增后降态势, 化肥减量成为当前农业碳减排的最大贡献者。中国农用氮、磷、钾化肥的产量由2015年的最高值7.43×107 t (折纯, 下同)降至2020年的5.50×107 t, 降幅达26.05%; 化肥施用量由2015年的最高值6.02×107 t降至2020年的5.25×107 t, 降幅达12.82%; 中国化肥的碳排放由2015年的3.35×108 t CO2 eq降至2020年的2.74×108 t CO2 eq, 降幅达18.21%。有机肥产量呈上升态势, 促进了农业固碳减排, 2020年有机肥产量达到1.56×107 t, 比2015年增长29.46%。科学施肥技术加快推广, 提高了肥料吸收利用率, 水稻、玉米、小麦三大粮食作物化肥利用率逐年提高, 2020年达40.20%, 比2015年提高5个百分点。目前, 我国肥料行业还存在一些问题: 1)化肥施用量依然偏高, 利用率偏低; 2)新型肥料创新不足, 市场较为混乱; 3)施肥不科学问题依然存在, 化学肥料偏多、有机肥偏少; 4)肥料立法缺失, 监管仍然薄弱。对标“双碳”目标, 我国肥料行业发展应该做好5个方面工作: 大力研发新型肥料, 加快推广减肥增效科学施肥技术, 强化农业废弃物资源化综合利用, 强化肥料行业立法监管, 加强科学低碳施肥宣传培训。
“双碳”目标为农业绿色发展提出了新要求, 特别是农业生产方式的绿色转型尤为迫切。文章采用文献梳理归纳法, 基于对农业绿色发展内涵及原则的分析, 剖析当前农业绿色发展现状及未来发展重点, 并进一步从生产、产业、经营和政策四大体系层面系统梳理农业绿色发展的研究进展; 最后, 对“双碳”目标下农业绿色发展研究现状进行了评述, 并对未来研究趋势进行了展望。研究表明: 农业绿色发展研究的内容主要聚焦于基础理论、指标测度、发展现状以及生产体系、支持政策等方面, 而对“双碳”目标下农业绿色发展的内涵与外延、体系创新等方面的研究较为薄弱。未来“双碳”目标下农业绿色发展研究应注重科学界定农业绿色发展的新内涵, 全面阐述“双碳”目标与农业绿色发展之间的关系, 健全具有中国本土化特色的农业绿色发展水平评价指标体系; 在此基础上, 从农业全产业链的视角诊断农业绿色发展问题, 对标“双碳”目标研判未来发展重点, 特别是探索多维度创新方向, 深度融入数字赋能、主体培育、市场引导、组织引领等创新要素, 从四大体系方面加快“双碳”目标下农业绿色发展体系创新的探究, 以期为农业绿色发展和全面推进乡村振兴战略提供科学依据和理论支撑。
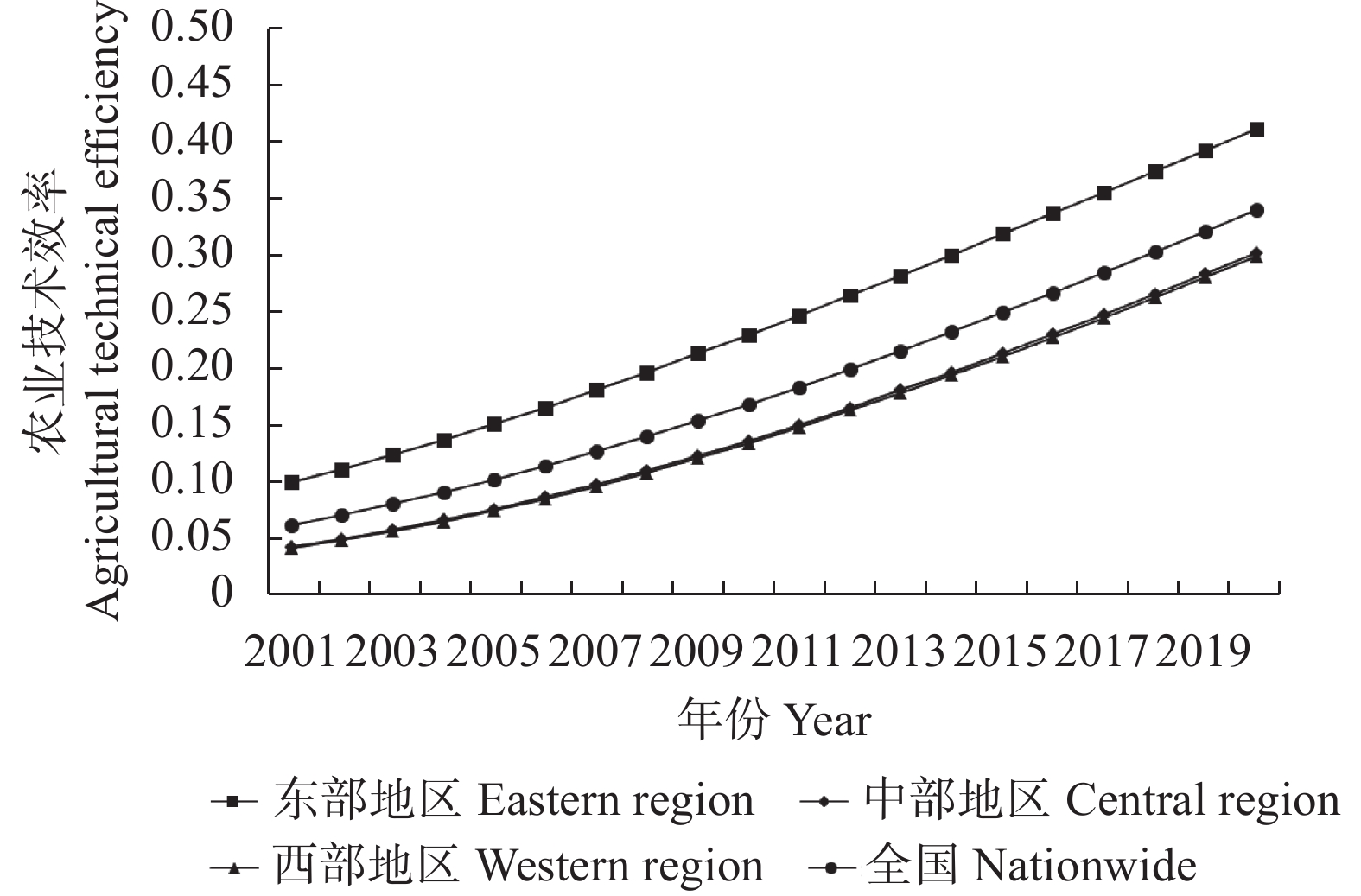
农业技术是促进农业产业发展的根本力量, 探究其对农业碳排放的影响机制, 对实现我国“双碳”目标具有重要意义。本文基于2001—2020年我国31个省、直辖市和自治区(港澳台地区以外)的面板数据, 使用随机前沿模型对农业技术效率进行测算, 并对各地区农业碳排放总量与强度进行核算, 构建空间杜宾模型和以农业技术效率为门槛变量的门槛模型, 探究农业技术效率和农业碳排放的空间效应与非线性关系。结果表明: 全国农业碳排放总量与强度近年来呈下降趋势。中部地区农业碳排放总量高于东西部地区, 东部地区农业技术效率高于中西部地区, 而农业碳排放强度则低于中西部地区。农业碳排放强度与农业技术效率具有空间自相关性, 并表现为集聚特征, 集聚类型以高高聚聚和低低聚集为主。农业碳排放强度具有正向空间溢出效应, 而农业技术效率对农业碳排放强度则表现为负向空间溢出, 此外城镇化、人力资本水平和人均耕地面积对农业碳排放强度具有负向影响, 农业经济发展水平和农业受灾程度为正向影响。农业技术效率与农业碳排放强度存在双门槛效应, 当农业技术效率达到“拐点”后, 其对农业碳排放强度的影响转变为负向, 当进一步提升农业技术效率水平后, 其影响力会因边际效应递减而减弱。本研究为探索实现“双碳”目标的路径提供理论基础与政策依据。
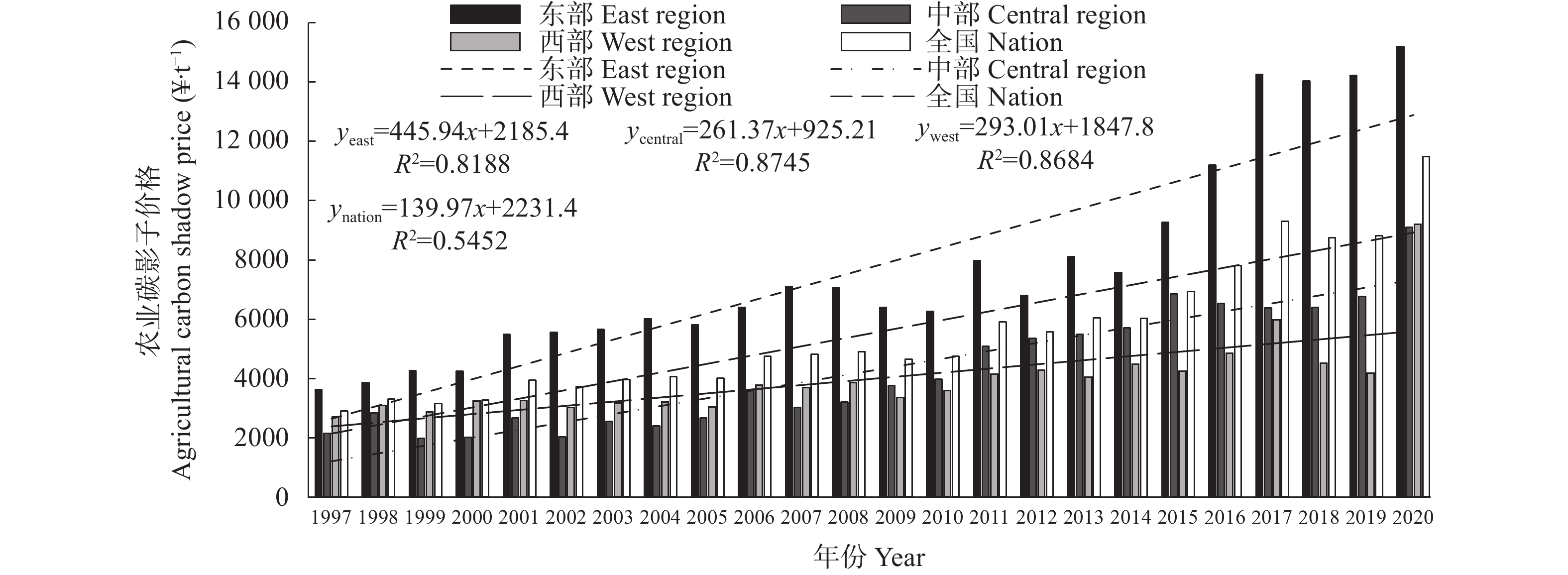
通过经济结构转型和技术进步有效实施减排策略, 已经成为实现中国经济社会低碳转型发展的必然选择。基于数据包络分析(DEA)框架建模、满足经济理论和物料守恒原则, 利用改进的By-production (BP)技术对1997—2020年我国31个省份农业碳影子价格进行了更加准确的测度, 并利用核密度分析了农业碳影子价格动态演变特征; 并采用可行广义最小二乘法(FGLS)模型考察结构转型、技术进步选择对农业碳排放影子价格的影响。结果表明: 1)我国农业碳影子价格呈上升态势, 东部、中部和西部地区农业碳排放影子价格分别为7759.69元∙t−1、4192.35元∙t−1和3997.51元∙t−1, 且东、中、西部地区农业碳影子价格上升趋势依次降低。2)我国农业碳影子价格核密度值有增加趋势; 东部地区农业碳影子价格核密度曲线出现较为明显的右移趋势; 中部地区农业碳影子价格核密度曲线呈现左移-右移趋势, 且区域间差异在变大; 西部地区农业碳影子价格核密度曲线呈现明显的向下、变宽趋势。3)整体回归显示, 结构转型、劳动节约型技术进步显著提升了农业碳影子价格, 而资本深化抑制了农业碳影子价格提升, 经济发展水平、农业经营规模、城市化水平及对外开放水平等对农业碳影子价格也会产生重要影响。4)东、中、西部地区部分样本回归显示农业碳影子价格影响因素有所差异, 结构转型在东部地区显著提升农业碳影子价格, 在西部地区却显著抑制了农业碳影子价格; 劳动节约型技术进步在东部抑制了农业碳影子价格, 而在西部显著提升农业碳影子价格; 资本深化在东、西部显著抑制农业碳影子价格, 在中部显著提升农业碳影子价格。为此, 提出持续推进产业结构转型、制定差异化绿色协调发展政策、适时建立农业碳排放交易市场等政策建议, 以促进农业低碳绿色高质量发展。

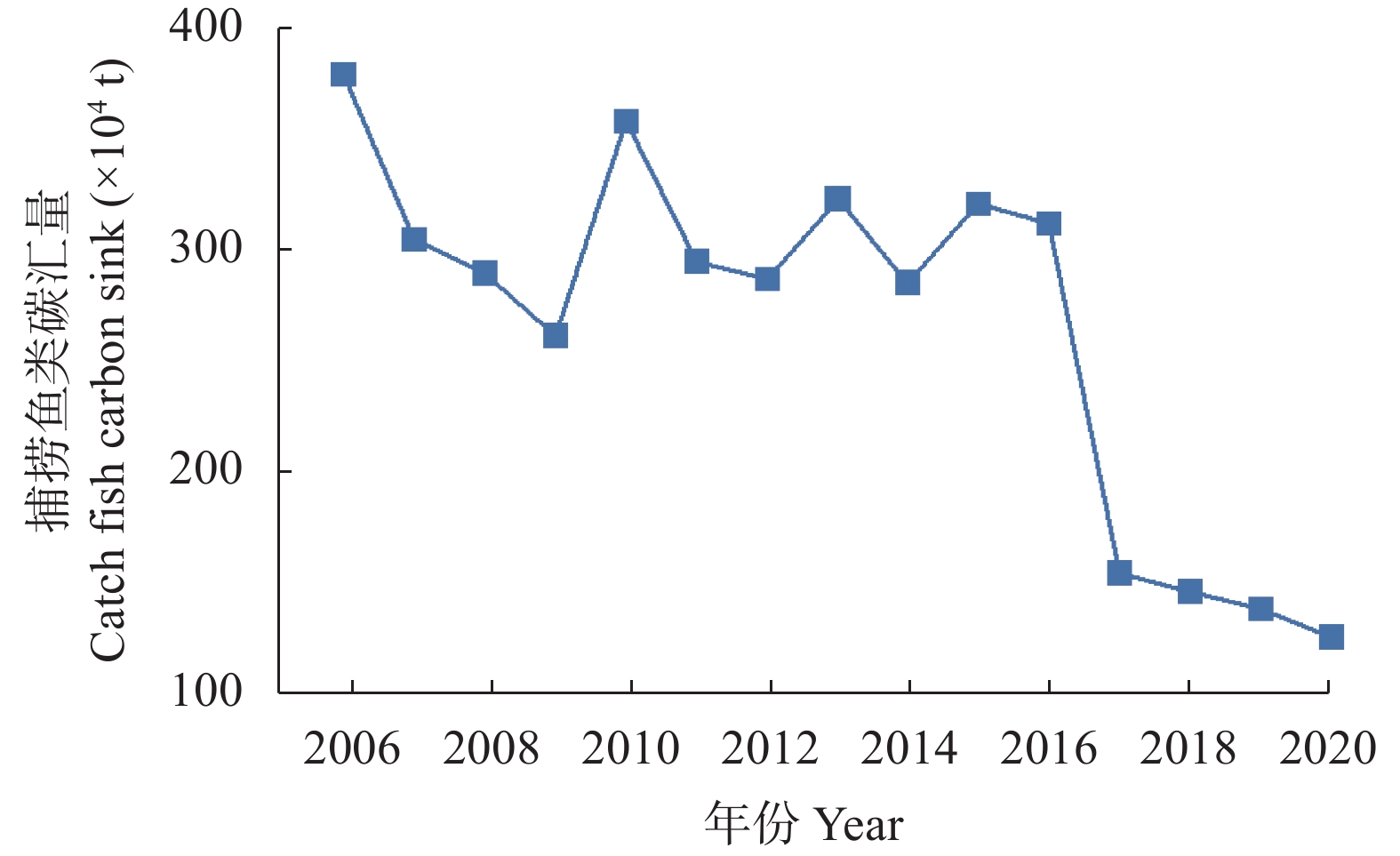
海洋渔业碳汇是海洋碳汇的主要组成部分, 是实现海洋碳增汇的有效途径之一。在碳达峰与碳中和背景下, 海洋渔业兼具“碳源”与“碳汇”的双重属性。利用《中国渔业统计年鉴》《国内机动渔船油价补助用油量测算参考标准》和《中国统计年鉴》数据, 计算了2006—2020年辽宁省渔业净碳汇量和渔业碳汇价值量; 运用时间序列三次指数平滑模型, 预测2021—2030年渔业碳汇量和渔业碳汇价值量; 基于灰色关联模型分析了辽宁省海洋渔业碳源碳汇量变化及其价值量变化的主要驱动要素。结果表明: 2006—2020年辽宁省海洋渔业碳汇收支盈余态势逐年减少, 海洋渔业碳汇赤字情况逐年加剧, 海洋渔业碳源碳汇最大顺差为256.36万t, 最大逆差29.99万t, 其平均差额为116.66万t·a−1。其中, 海洋捕捞鱼类碳汇总量3976.04万t, 但自2016年起急剧下降, 并呈持续下降趋势; 贝藻类碳汇总量241.67万t, 养殖占83%, 变化不大; 海洋捕捞碳排放量为164.52万t·a−1, 其中拖网捕捞渔业占比近50%。2017年后海洋捕捞碳汇量不能补偿碳排放量。辽宁省海洋渔业碳汇价值总量274.23亿元, 年均18.28亿元。辽宁省渔业碳汇总量和渔业碳汇价值总量持续下降, 碳汇价值量与碳汇量呈正相关。海洋渔业碳汇量与海洋捕捞渔获物产量、养殖贝藻类产量呈正相关。基于时间序列预测模型分析显示, 2020—2030年辽宁省海洋渔业碳汇赤字将持续加剧, 海洋渔业碳汇量逐年降低。辽宁省海洋渔业碳汇受国家政策、捕捞渔获物产量、从业人员数量、贝藻类养殖面积和海洋捕捞渔船总功率等因素影响。辽宁省海洋渔业碳源排放量受海洋捕捞渔船总功率、渔业专业户数量和技术推广机构数量影响明显。建议多种养殖方式深度融合, 减少高能耗、低产量捕捞作业方式, 保护海洋生物多样性, 并加强高排放渔船监管, 以促进辽宁省海洋渔业发展。
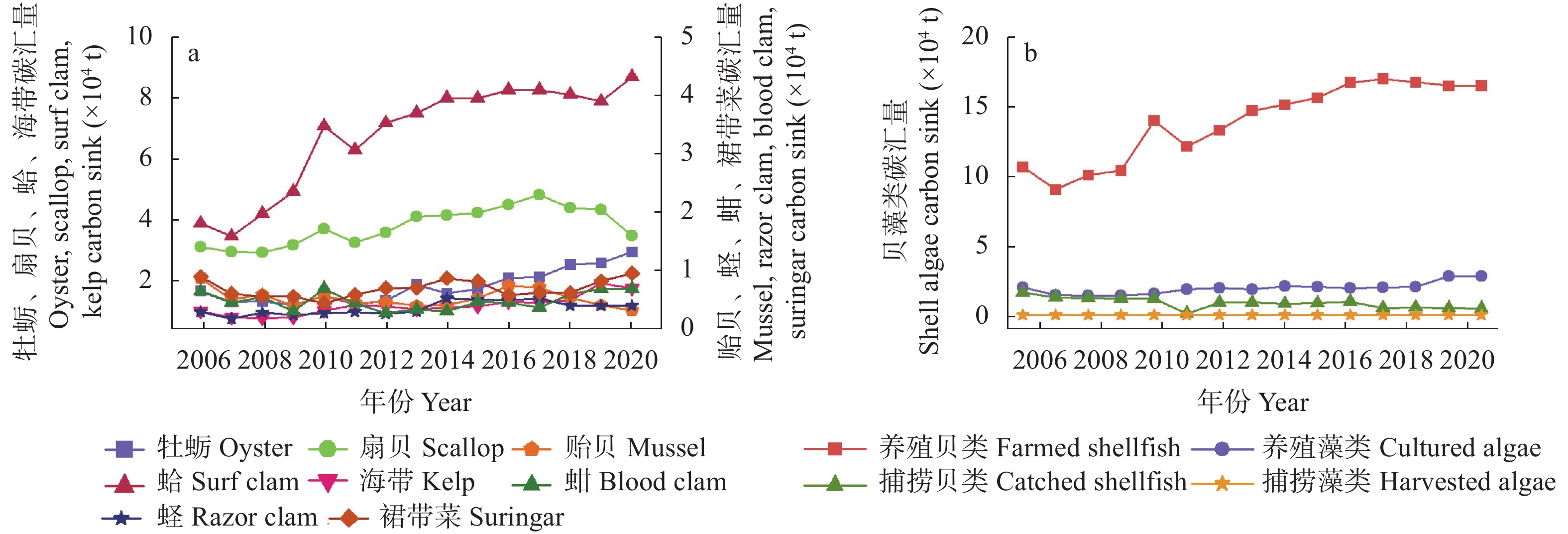
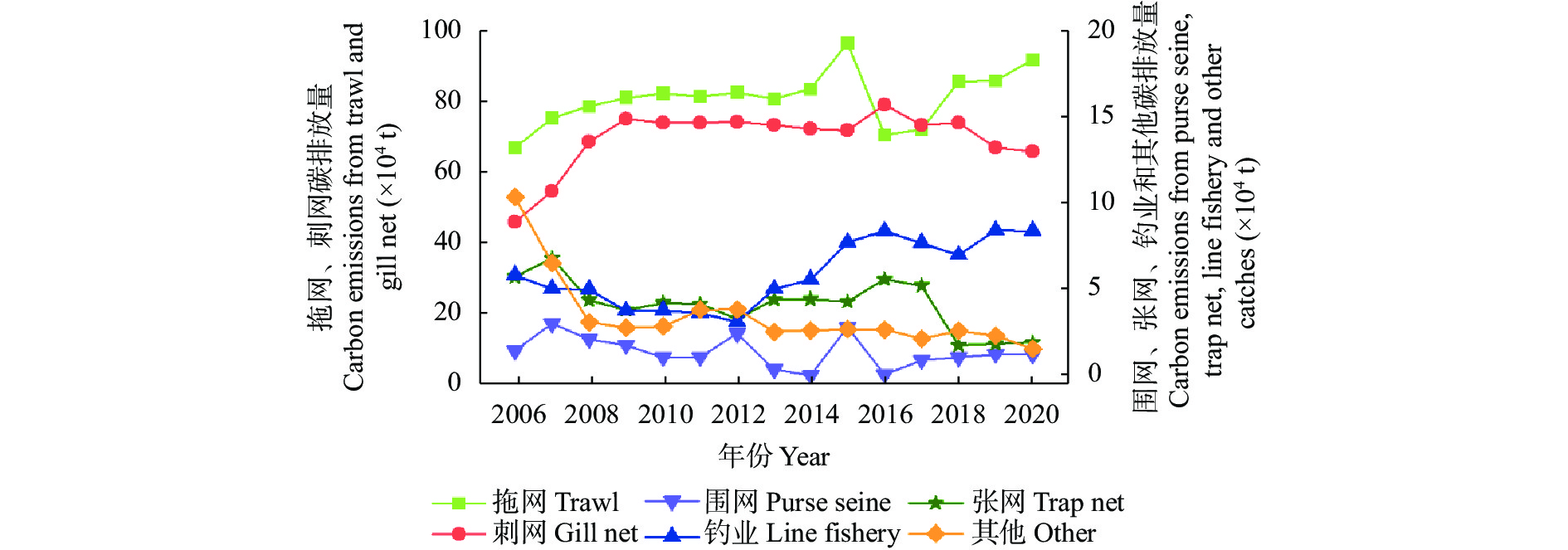
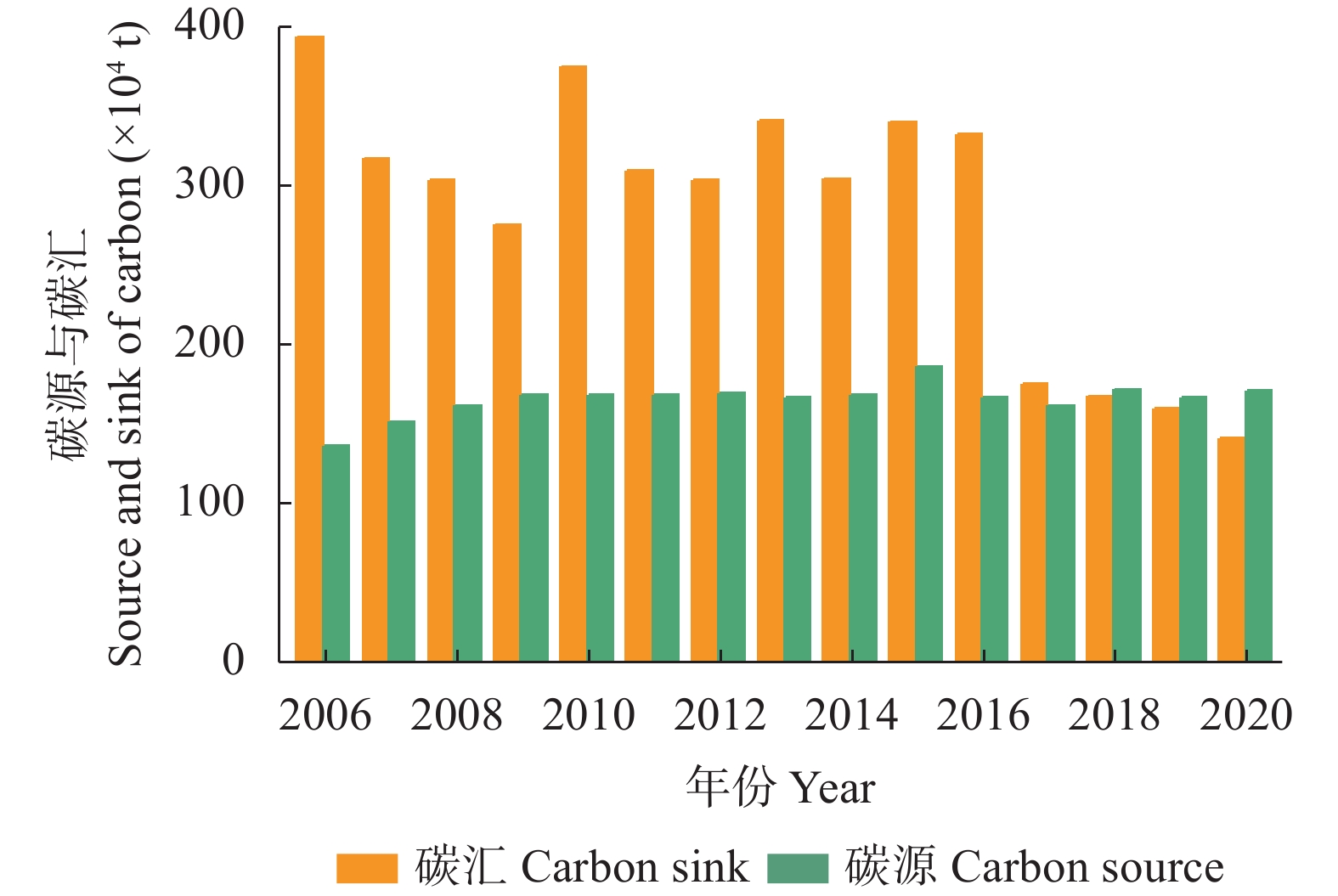

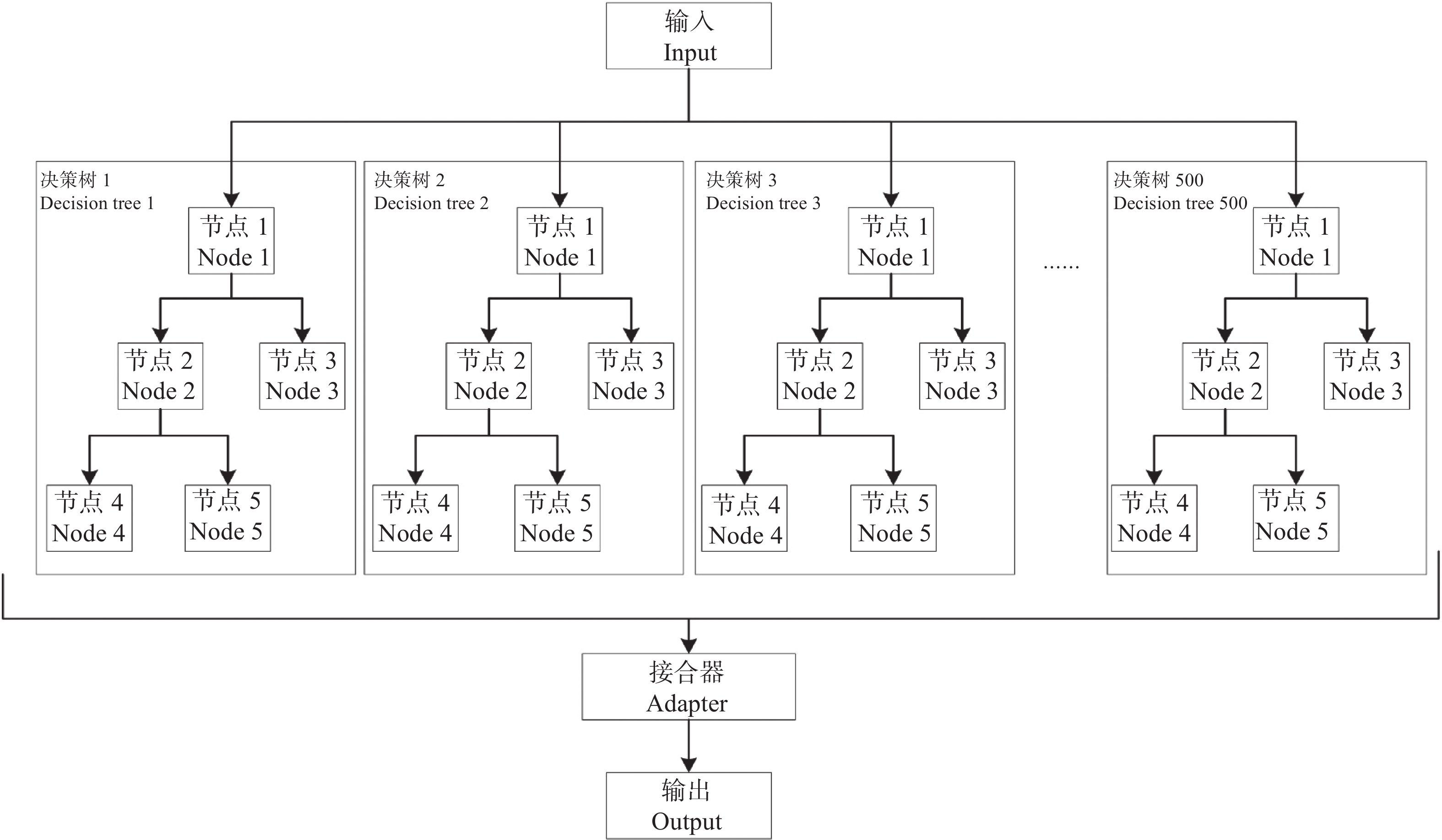
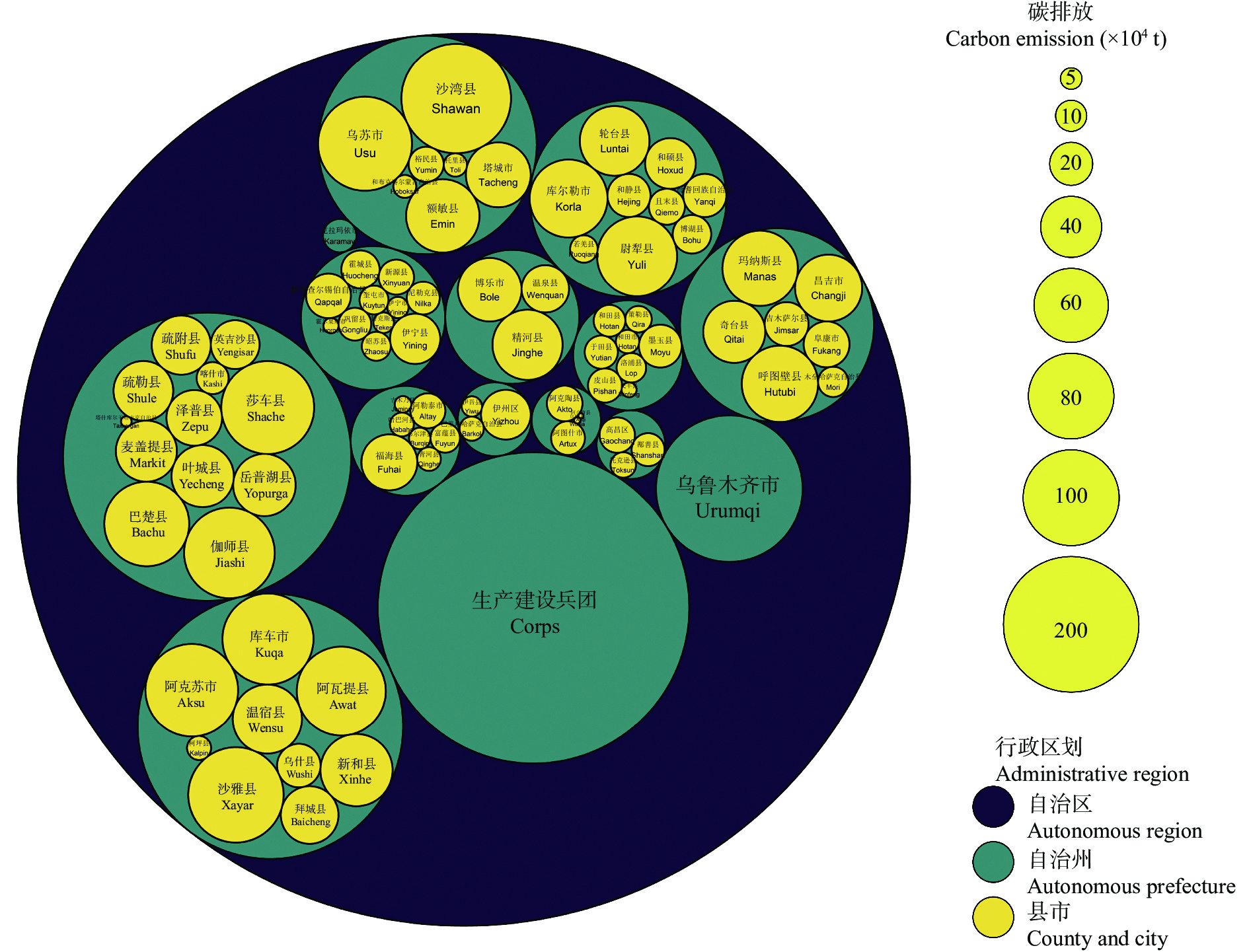
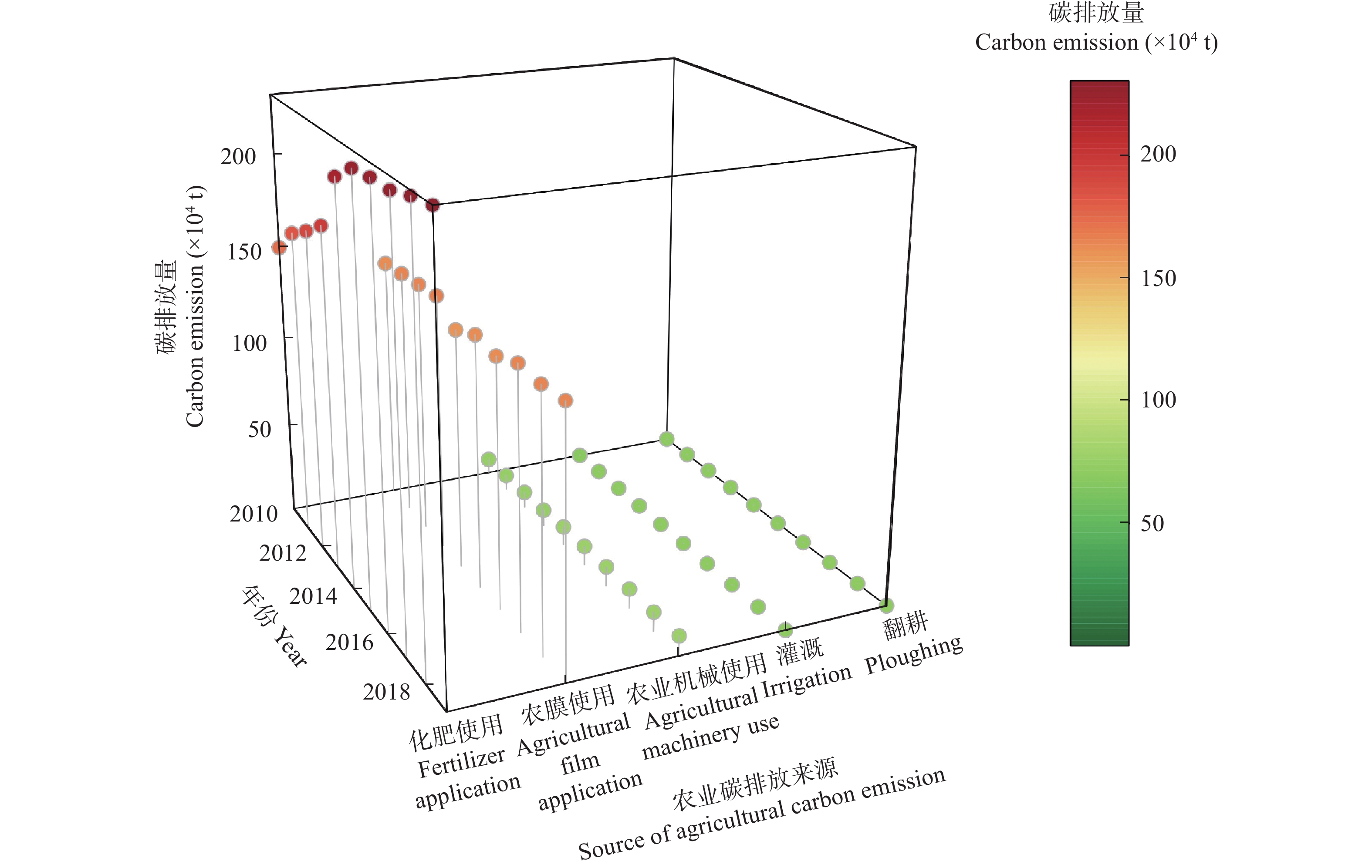
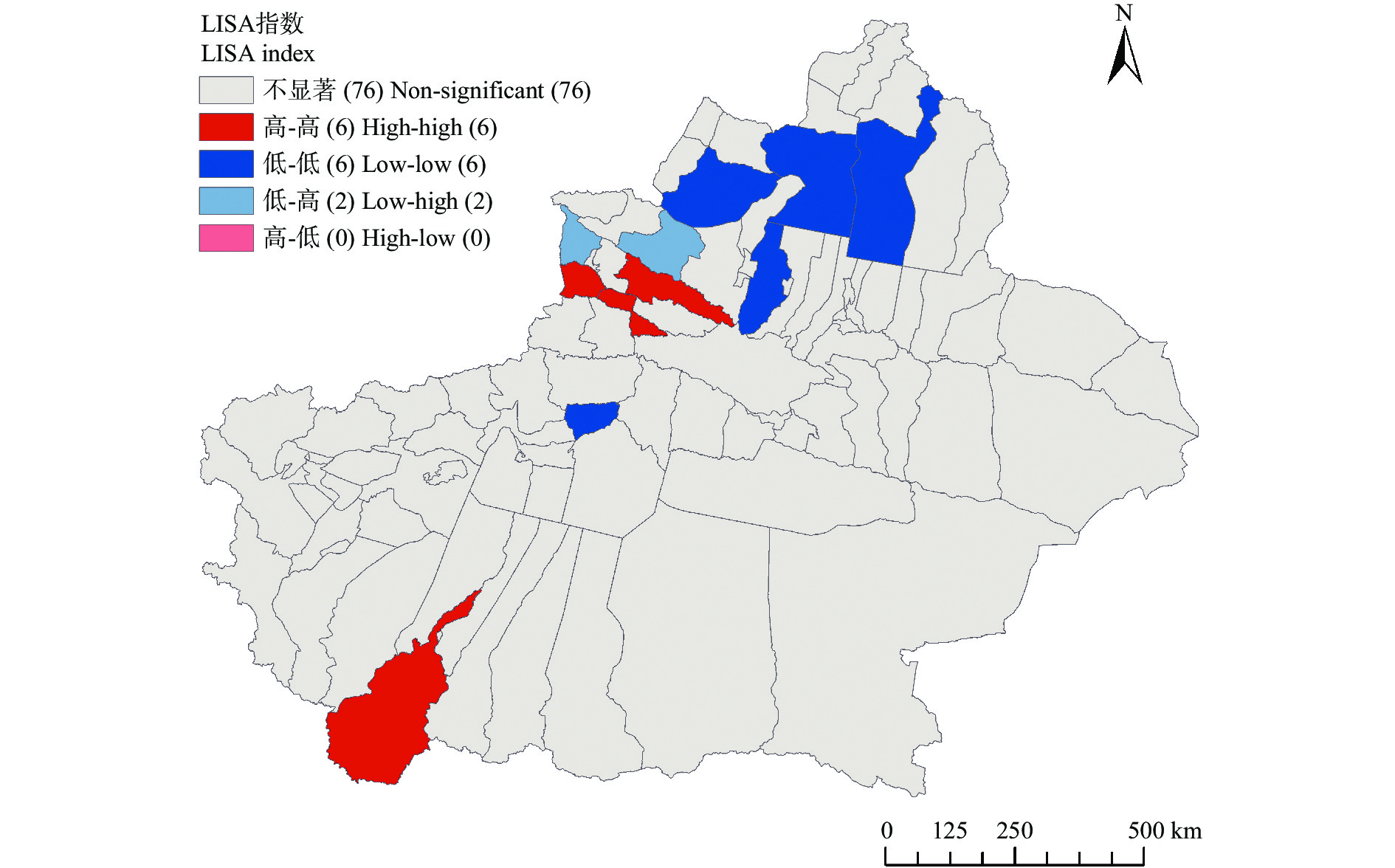
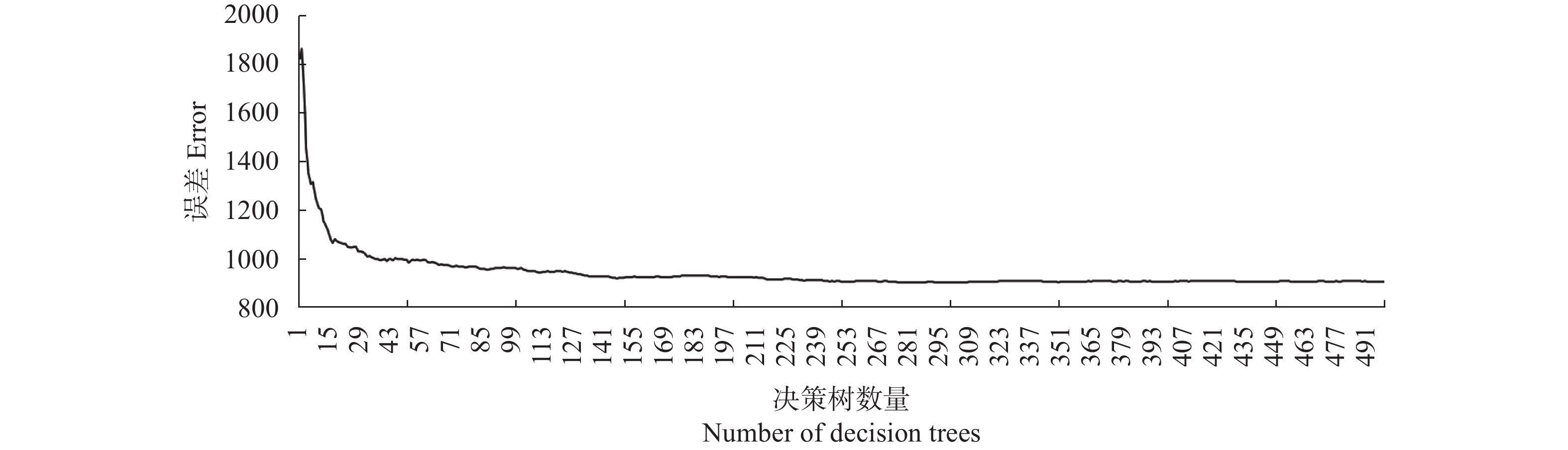
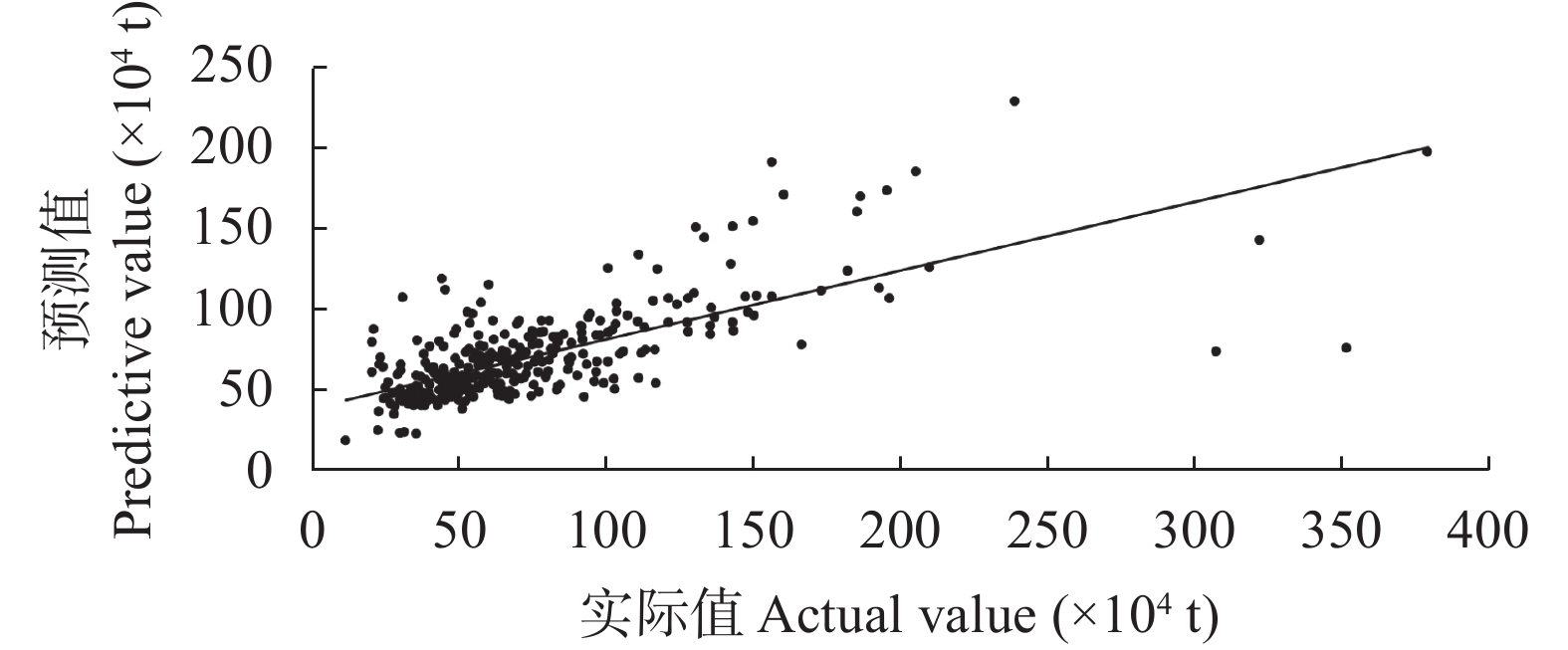
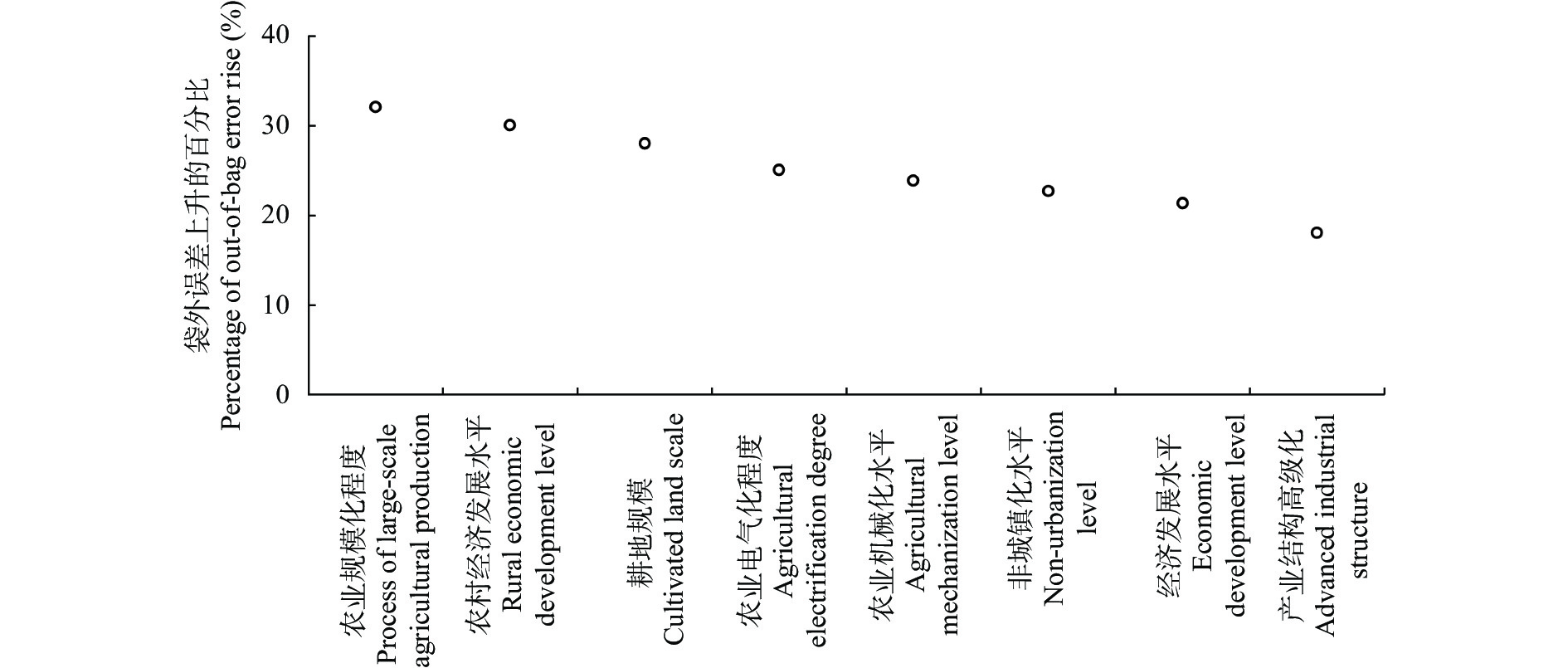
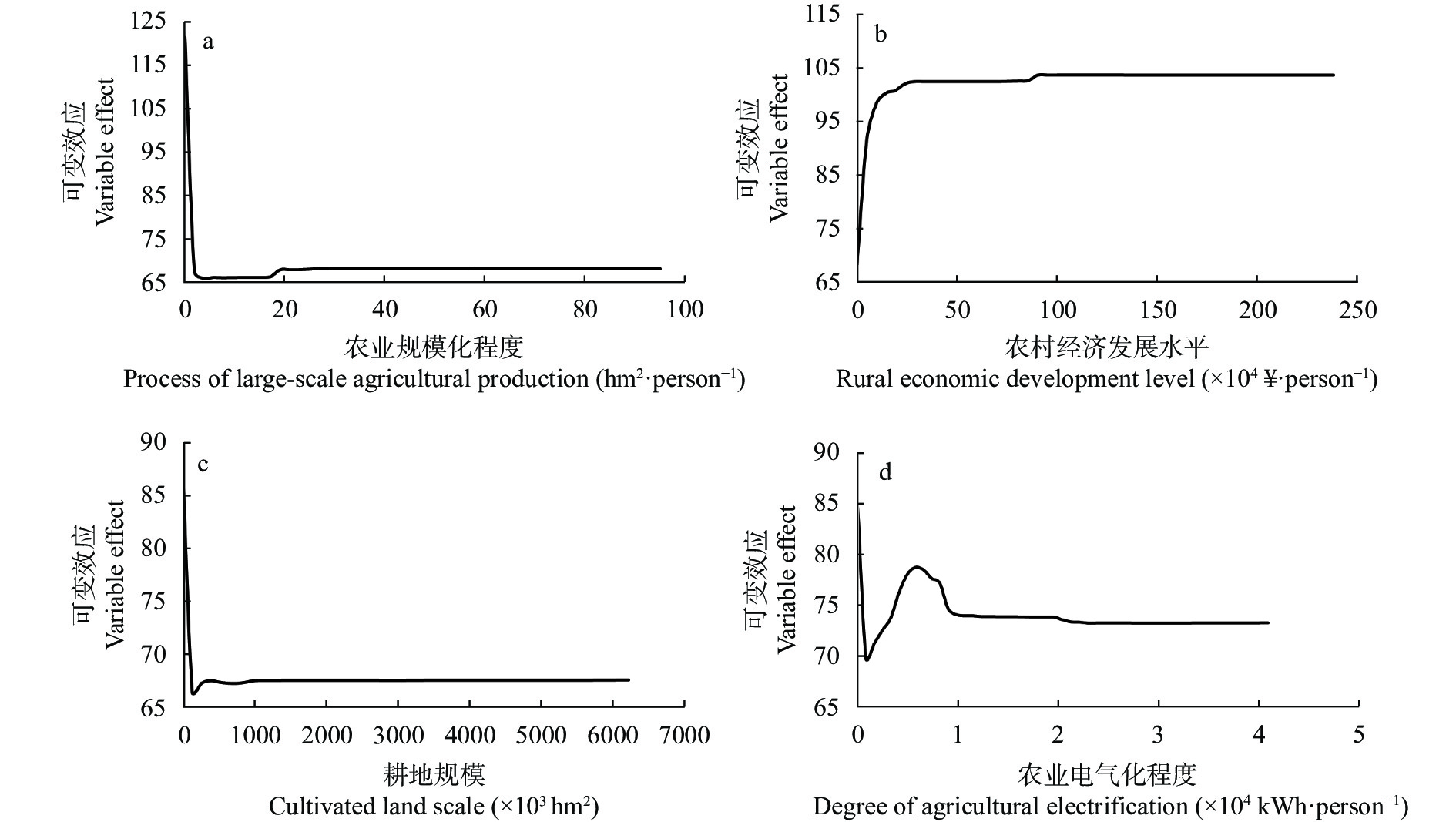
农业是全球第二大碳源, 明确农业碳排放规律对于碳达峰、碳中和具有重要意义。为探究新疆农业碳排放规律, 促进农业碳减排, 本研究根据农业生产过程中的碳排放环节, 结合国内外发布的碳排放系数, 测算了新疆的农业碳排放量; 利用莫兰指数、LISA指数等空间相关性模型测算了新疆农业碳排放的空间集聚规律; 利用机器学习中的随机森林模型对农业碳排效率影响因素进行了动态量化分析。结果显示: 1) 2010—2019年新疆农业碳排放量缓慢增长, 从292.24万t增长到379.69万t, 年均增速3.33%。2)化肥和农膜的使用是新疆农业碳排放的主要来源, 占比分别为58.06%和39.03%。3)新疆农业碳排放效率在不断提升, 2010—2013年增速较快, 2014—2019年增速较慢, 碳排放效率的主要分布区间从小于50元∙t−1变为50~100元∙t−1。4)新疆农业碳排放效率高高聚集区域农业产值不高, 主要是由于物质投入低; 低低聚集区域农业产值相对较高, 但科技、管理水平低, 物质投入过多。5)降水量较低的南疆区域, 农业碳排放效率整体较高, 降水量较高的北疆区域, 农业碳排放效率处于中等水平。6)农业规模化程度在0.12~2.02 hm2∙人−1时, 碳排放效率随着农业规模化程度提高急剧降低, 当农业规模化程度高于2.02 hm2∙人−1时, 对农业碳排放效率的影响力降低; 耕地规模在120~17 220 hm2时, 对农业碳排放效率有一个显著的负向影响, 当耕地规模大于17 220 hm2时, 对农业碳排放效率的影响较为平缓。农村经济发展水平对碳排放效率具有正向影响, 农业电器化程度对碳排放效率呈现出正“U”型影响。


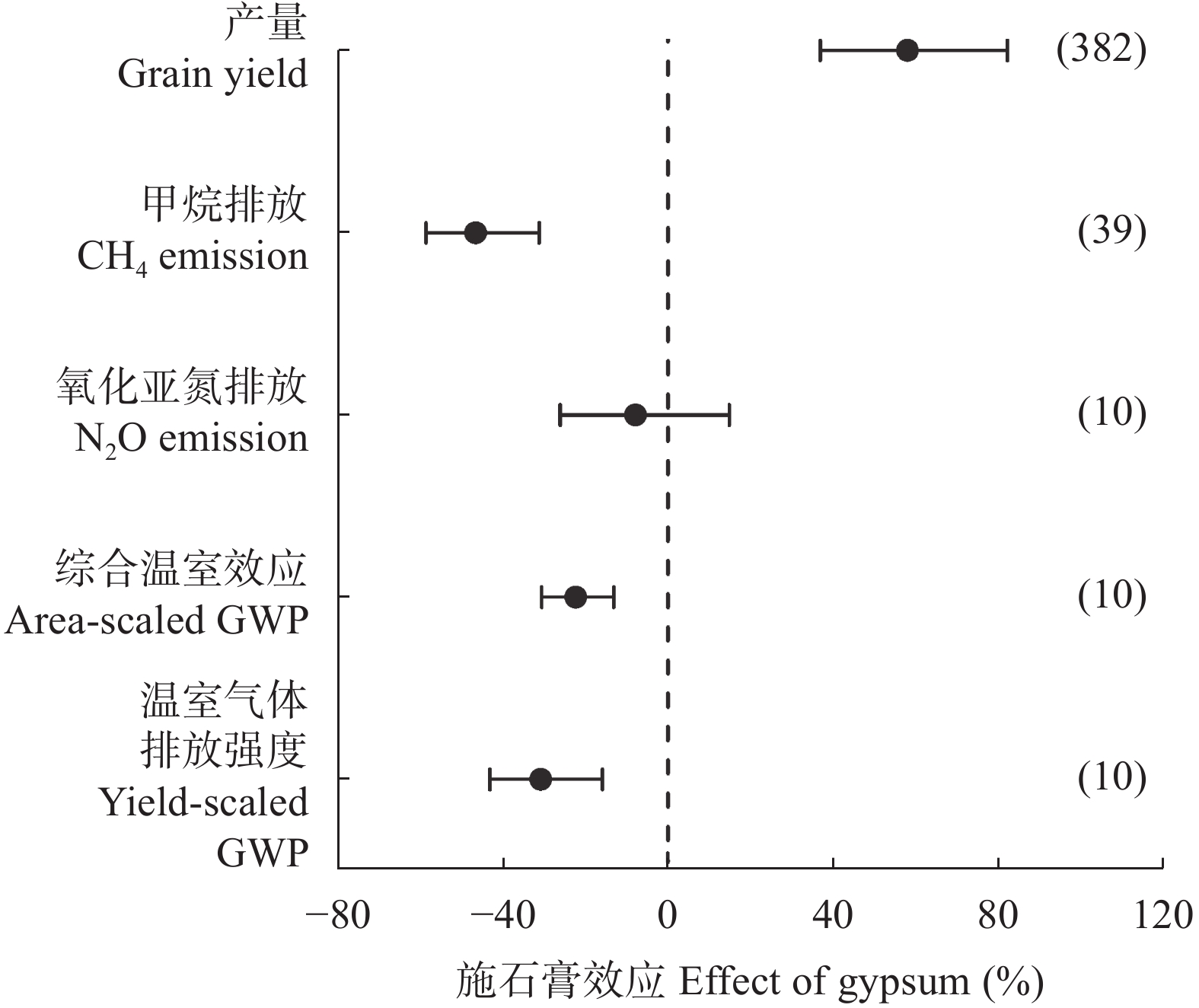
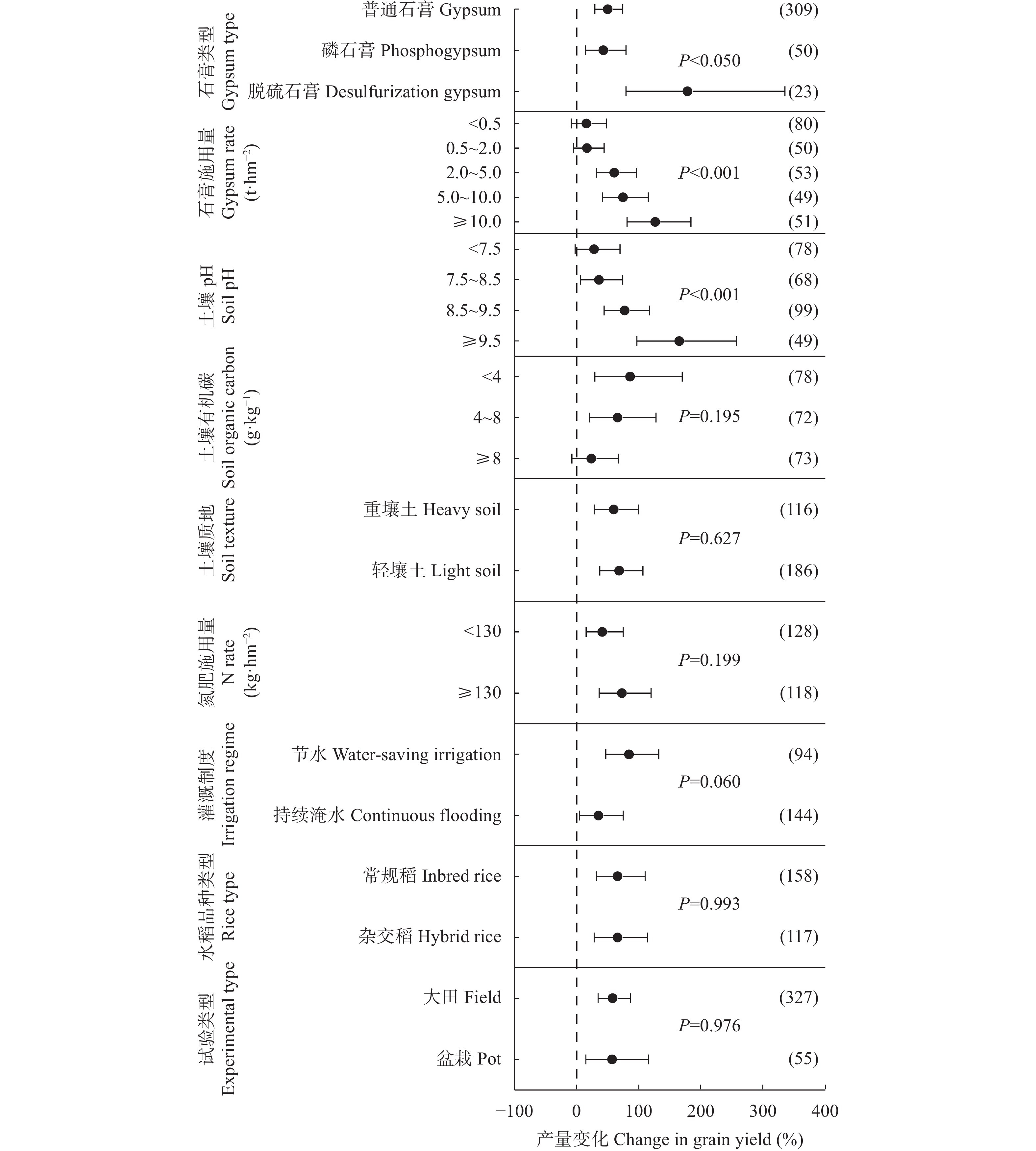
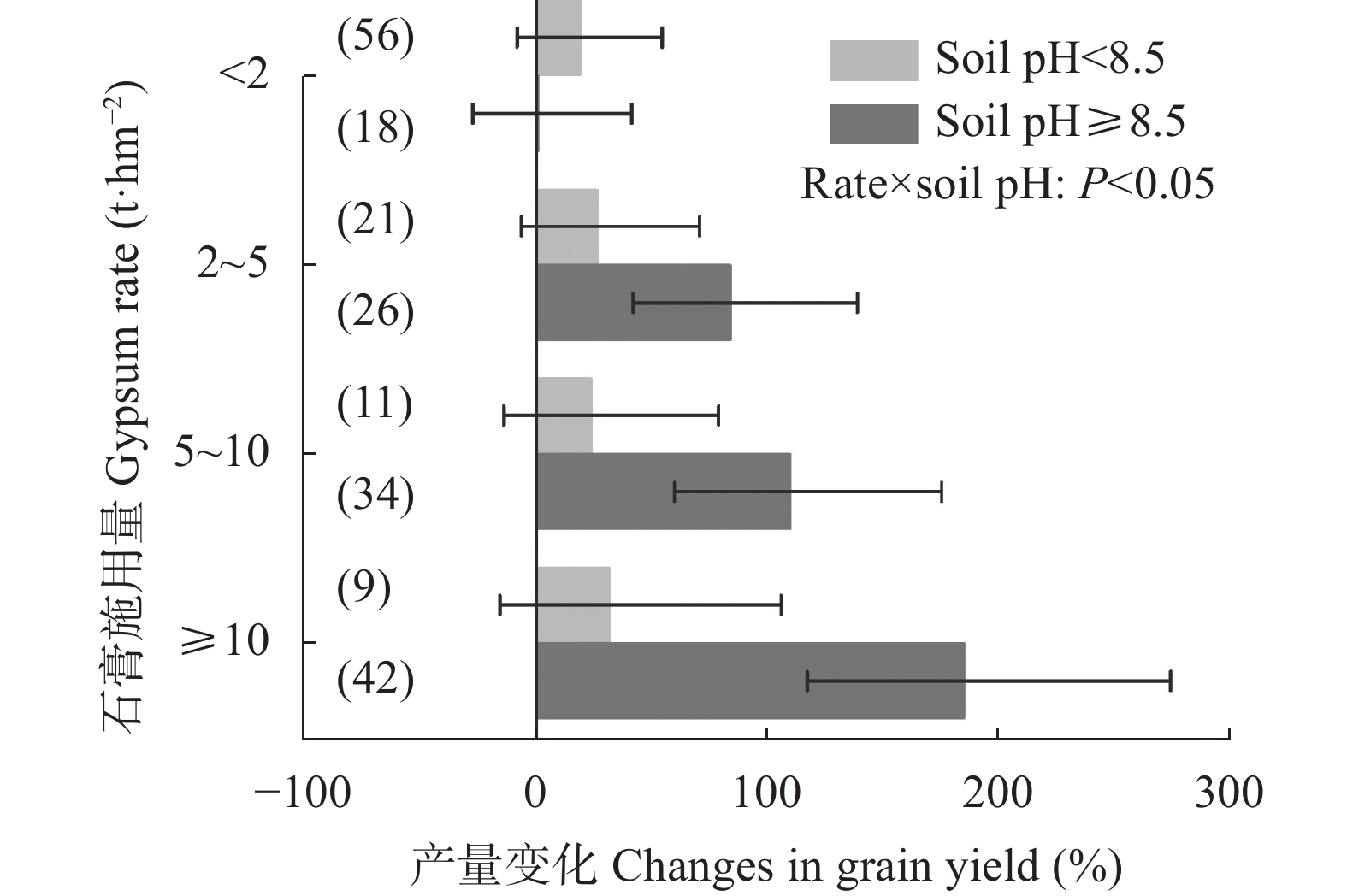
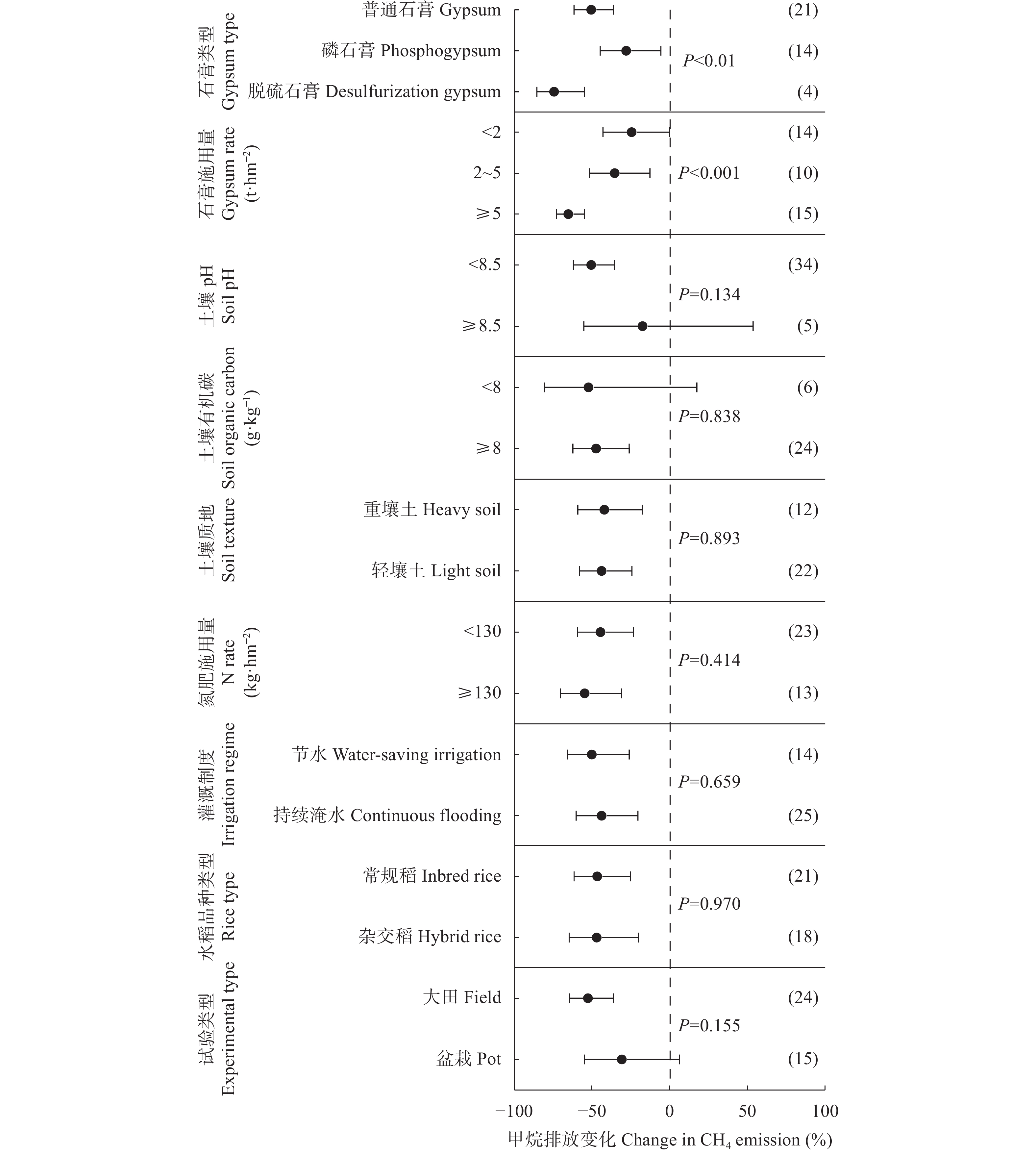
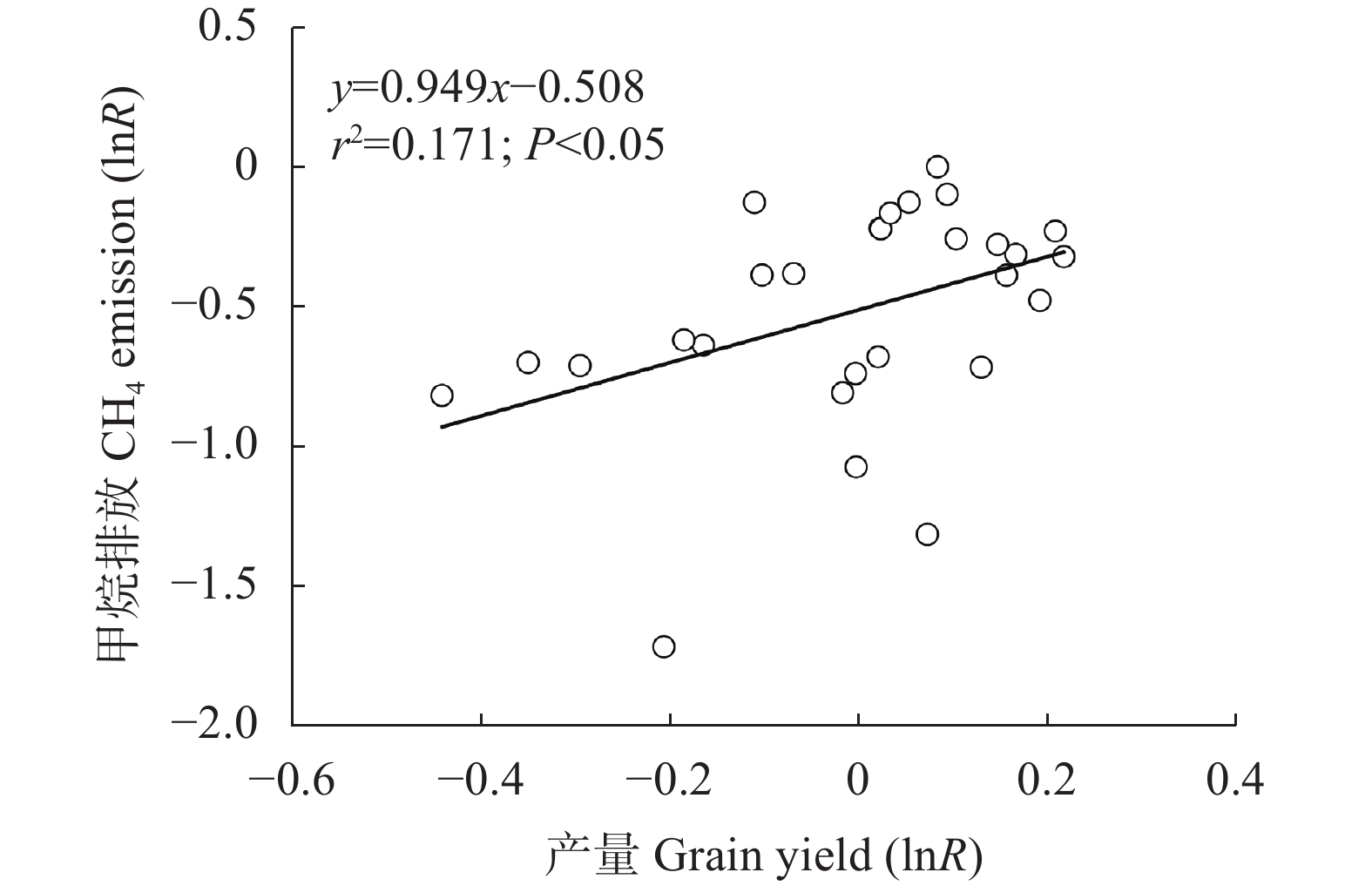
石膏是一种常见的稻田土壤改良剂, 施石膏对水稻产量和稻田温室气体排放影响的荟萃分析尚鲜见报道。本研究采用Meta分析方法, 探究施石膏对水稻产量和稻田温室气体排放的影响。以不施石膏为对照, 施石膏为处理, 在全球尺度上筛选出了74篇文献, 建立了包含382对水稻产量、39对甲烷(CH4)排放、10对氧化亚氮(N2O)排放、10对综合温室效应(GWP)和10对温室气体排放强度(GHGI)观测值的数据库。针对不同的石膏施用措施(类型和施用量)、基础土壤性状(pH值、有机碳含量和质地)以及稻田管理方式(氮肥施用量、灌溉制度、水稻品种类型和试验类型), 探究施石膏对水稻产量和稻田CH4排放的影响。从总效应来看, 与不施石膏相比, 施石膏显著增加了水稻产量(+58%), 降低了稻田CH4排放(−47%)、GWP(−22%)和GHGI(−31%), 而对N2O排放影响不显著。脱硫石膏对水稻增产和稻田CH4减排的效应显著高于普通石膏和磷石膏。当施用量<2 t·hm−2时, 石膏对水稻产量影响不显著; 当施用量≥2 t·hm−2时, 石膏对水稻的增产效应随石膏施用量的增加而增加。随着土壤pH增加, 施石膏对水稻产量的增幅显著增加。石膏施用量和土壤pH对水稻产量存在显著的互作效应。在土壤pH<8.5条件下, 施石膏对水稻产量影响不显著; 在土壤pH≥8.5条件下, 水稻产量随着石膏施用量的增加而增加。稻田CH4减排效应随石膏施用量的增加而显著增加。综上, 施石膏显著提高了水稻产量, 同时降低了稻田温室气体排放, 本研究结果可为评估施石膏对全球水稻丰产和缓解气候变暖提供数据支撑。
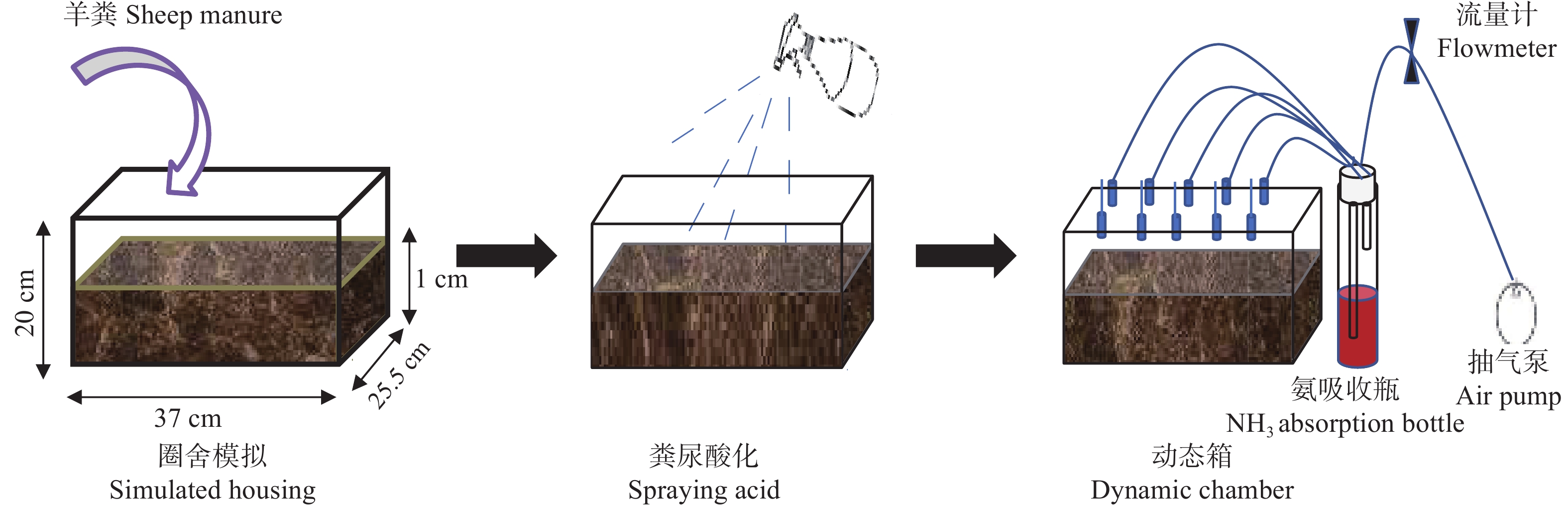
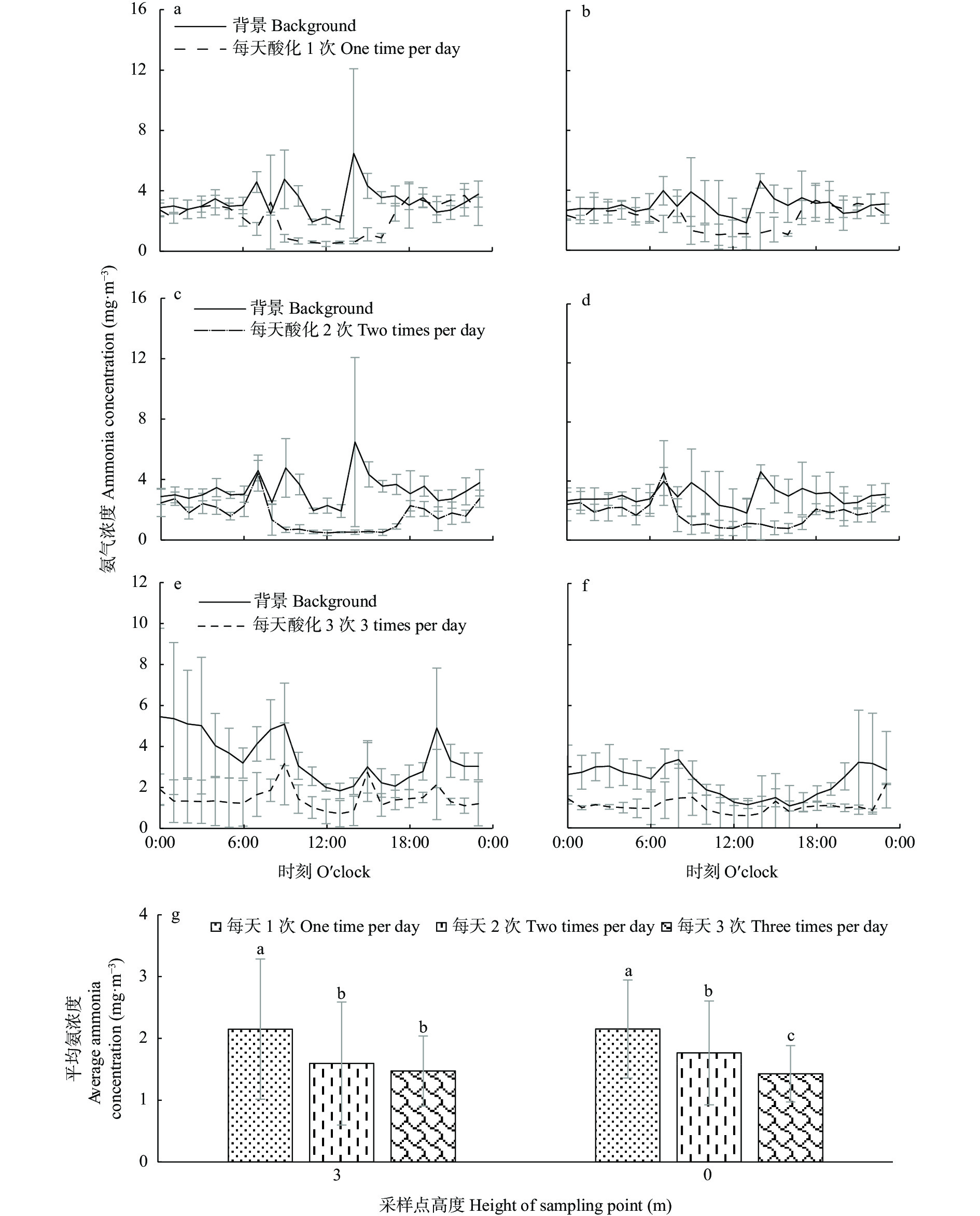
本研究针对养殖场圈舍环节氨挥发量大, 缺少可原位实施的氨减排技术的现状, 欲探究一种经济、高效、可实施并可广泛推广的圈舍原位氨减排技术。本研究利用圈舍模拟试验探索了不同剂量的硫酸和青贮渗出液对粪尿氨挥发的影响, 筛选可用于圈舍原位氨减排的酸化条件; 并进一步研发动物圈舍原位酸化氨减排设备, 将模拟试验筛选出的酸化条件应用于原位圈舍, 从氨减排效果和经济两个方面综合探究动物圈舍原位酸化技术的可行性。研究结果表明, 硫酸和青贮液可快速降低粪尿氨挥发: 以不喷酸作为对照, 按0.03 g∙m−2的喷施量, 在试验开始喷洒一次硫酸和青贮液, 可分别降低氨挥发排放速率39.1% (P<0.05)和42.7% (P<0.05), 但其减氨效果仅维持8 h左右。鉴于硫酸难以市场流通、青贮液无法雾化, 原位圈舍采用青贮液的主要成分乳酸作为酸化剂。本研究利用原位酸化氨减排设备对圈舍粪尿以不同频率喷酸时, 每天按0.03 g∙m−2的剂量喷洒乳酸(0.01 mol∙L−1) 3次(8:00、16:00、0:00)时氨减排效率最高, 与不喷酸相比, 氨减排效率为55.6% (P<0.01); 每天喷洒2次(8:00、16:00) 氨减排成本最低, 为147元∙kg−1(NH3)。本研究为养殖场圈舍环节提供了一项可实施的高效减氨技术, 但其成本仍较高, 该技术的广泛推广和应用还需进一步降低设备成本。
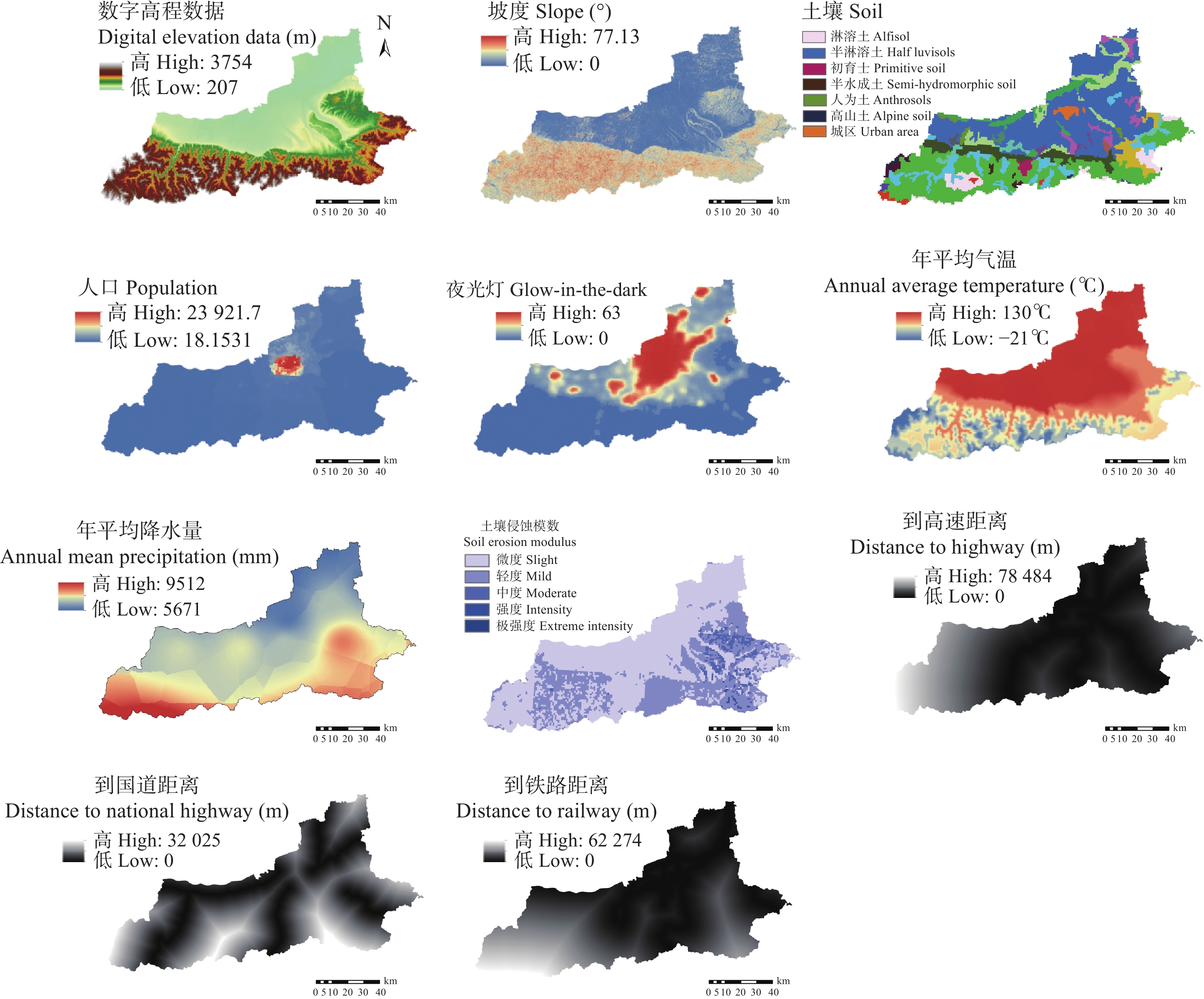
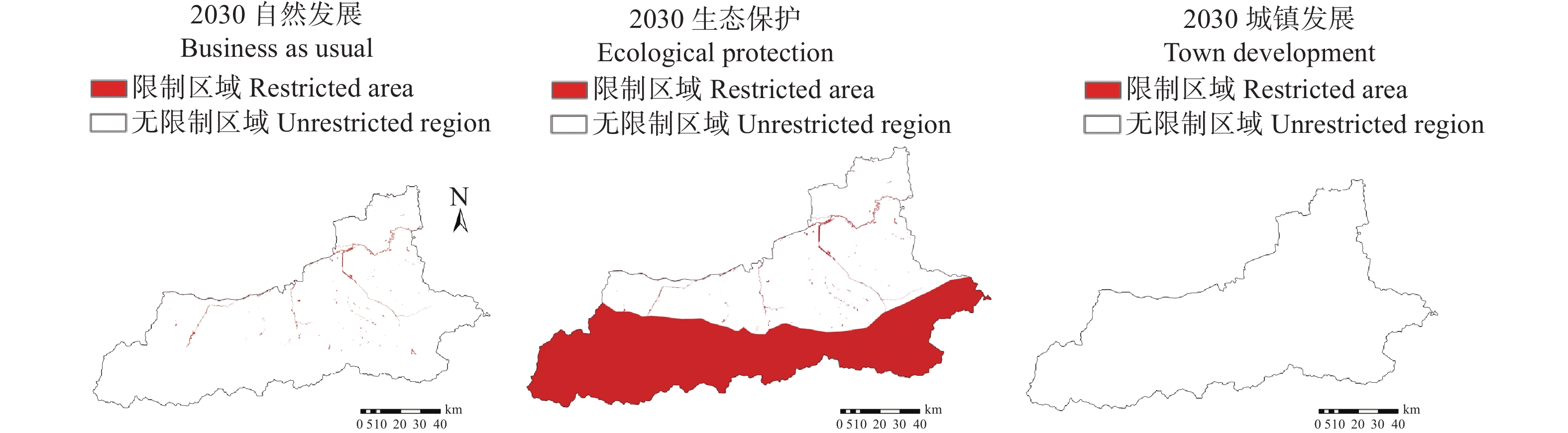
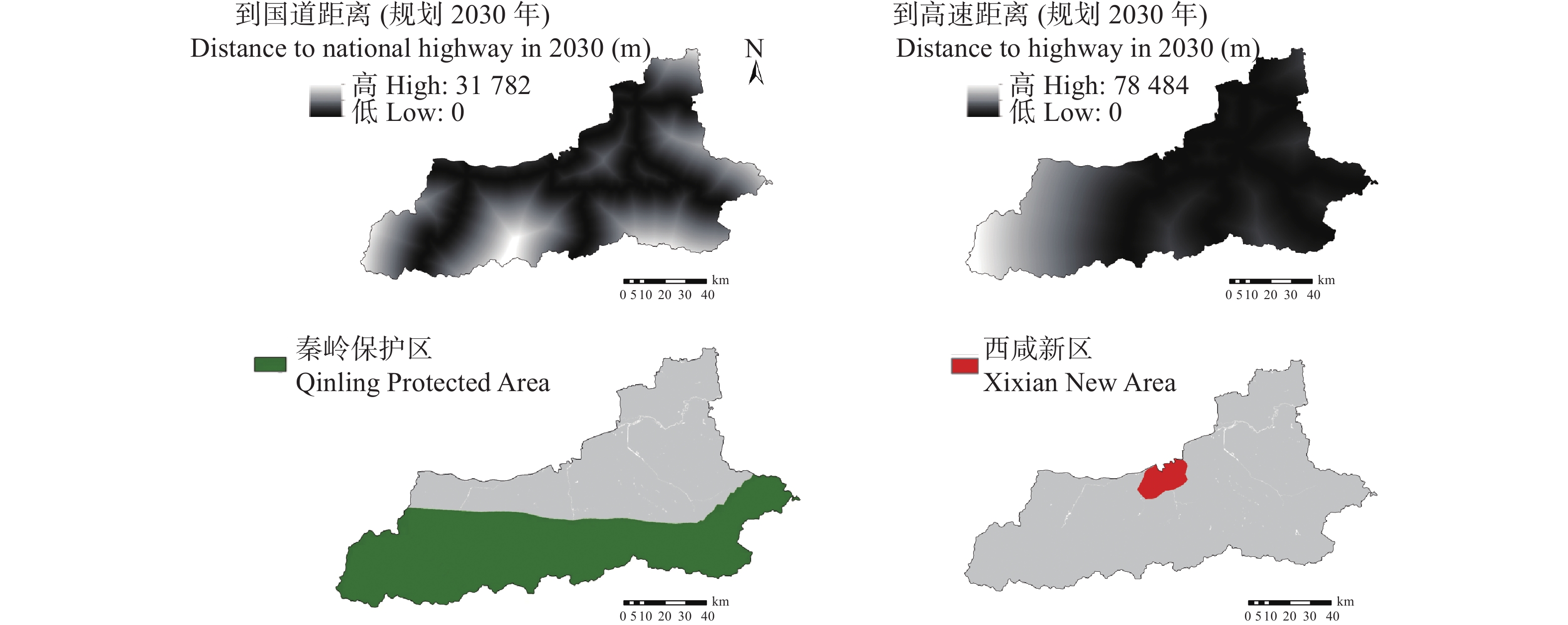
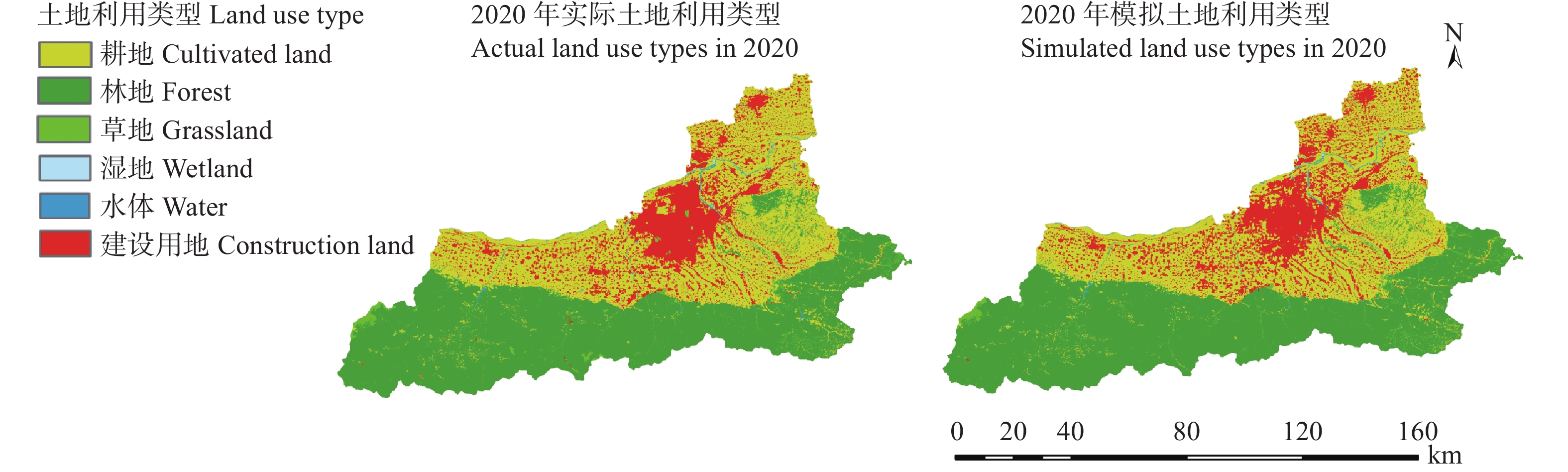
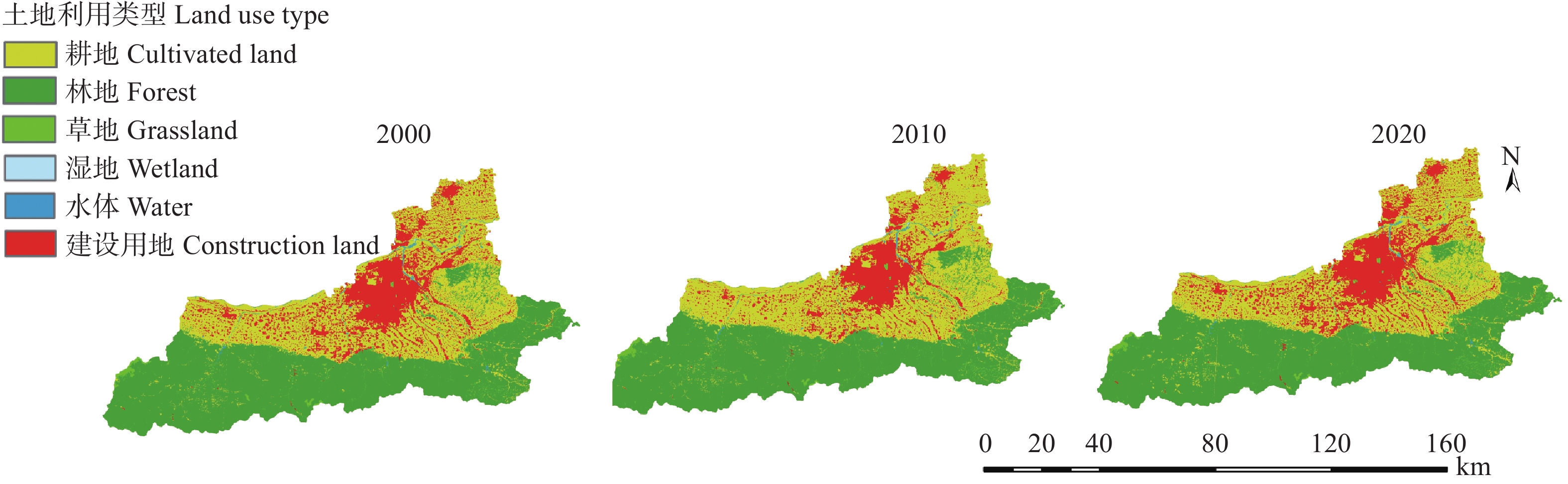
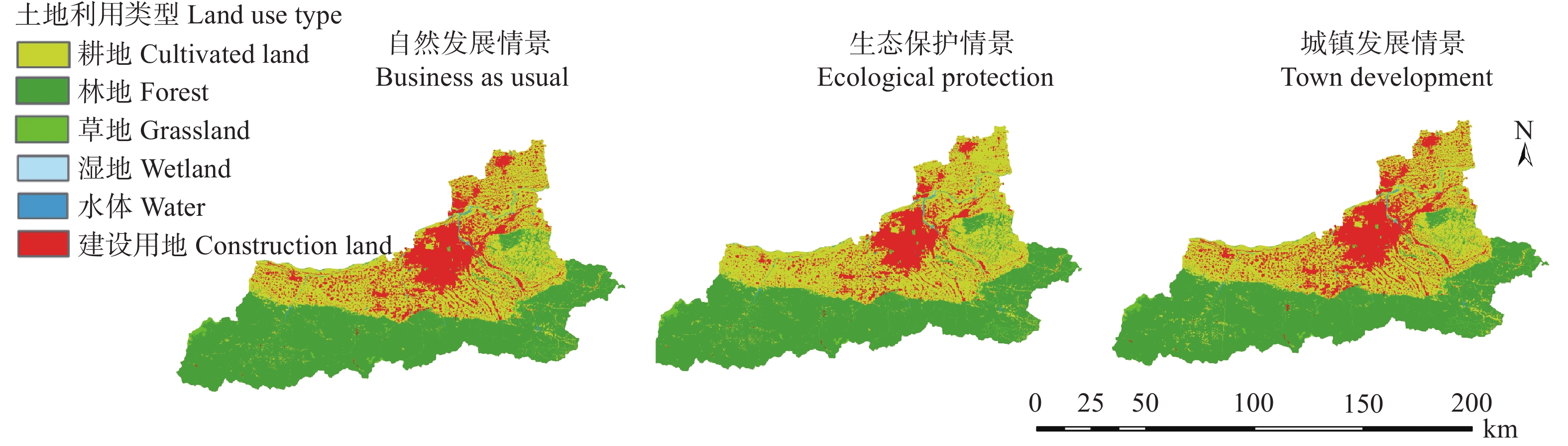
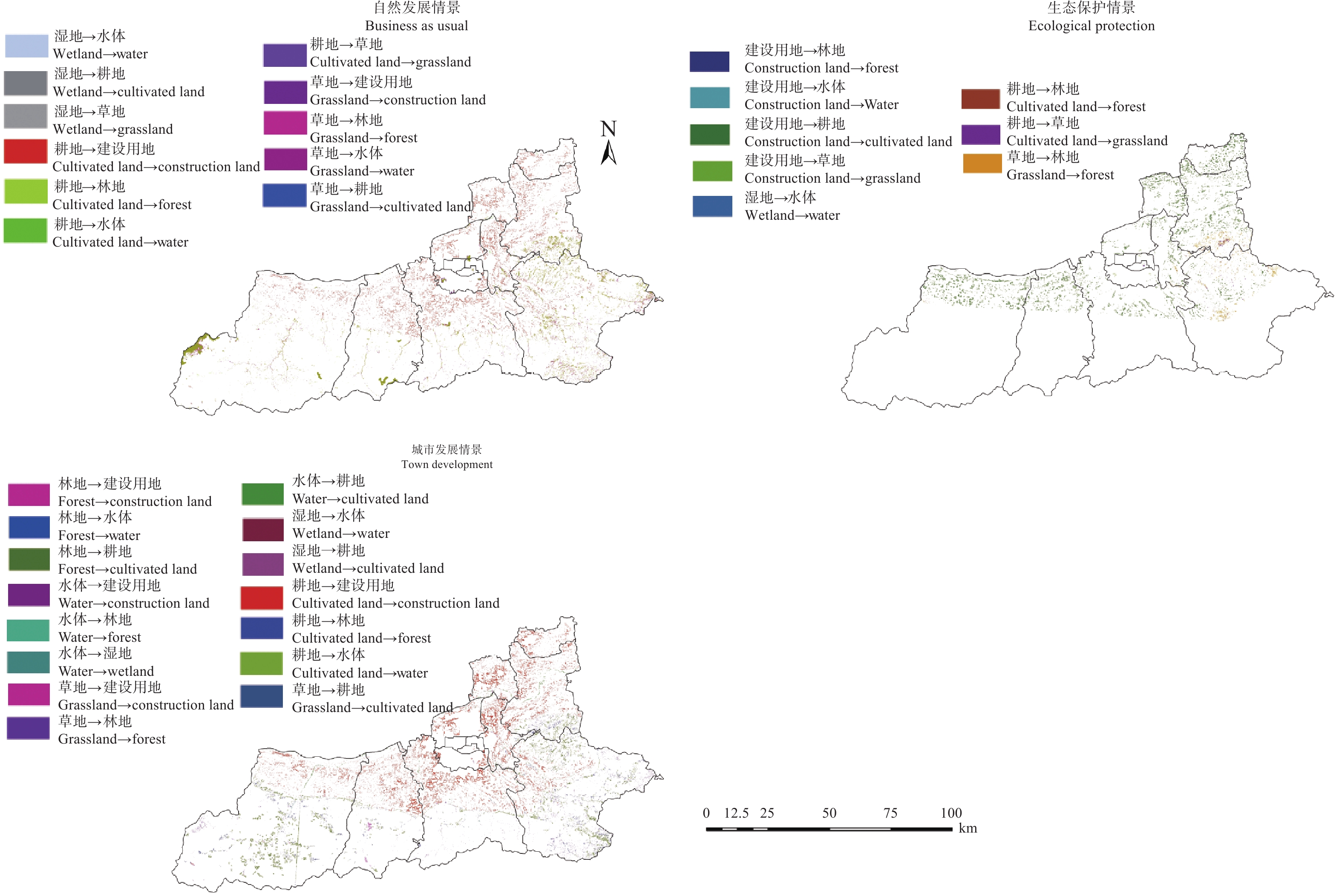
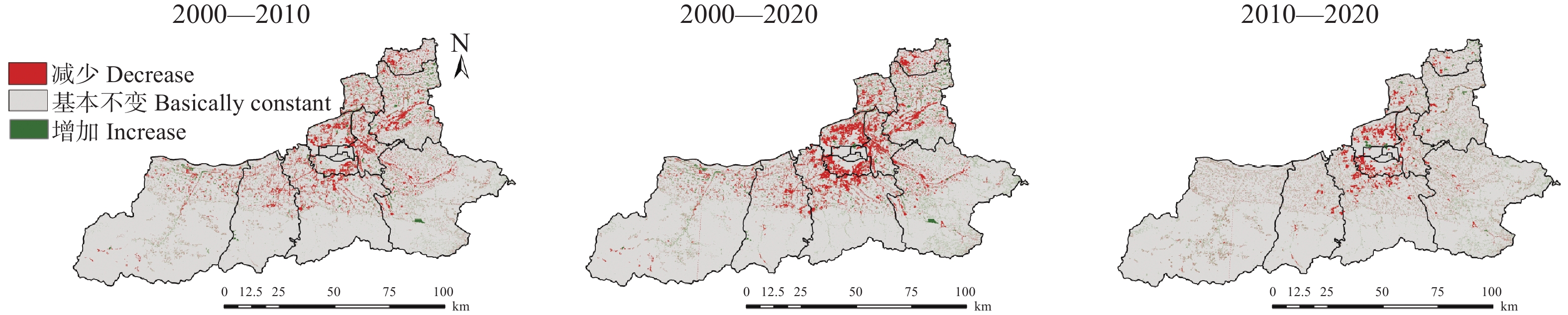
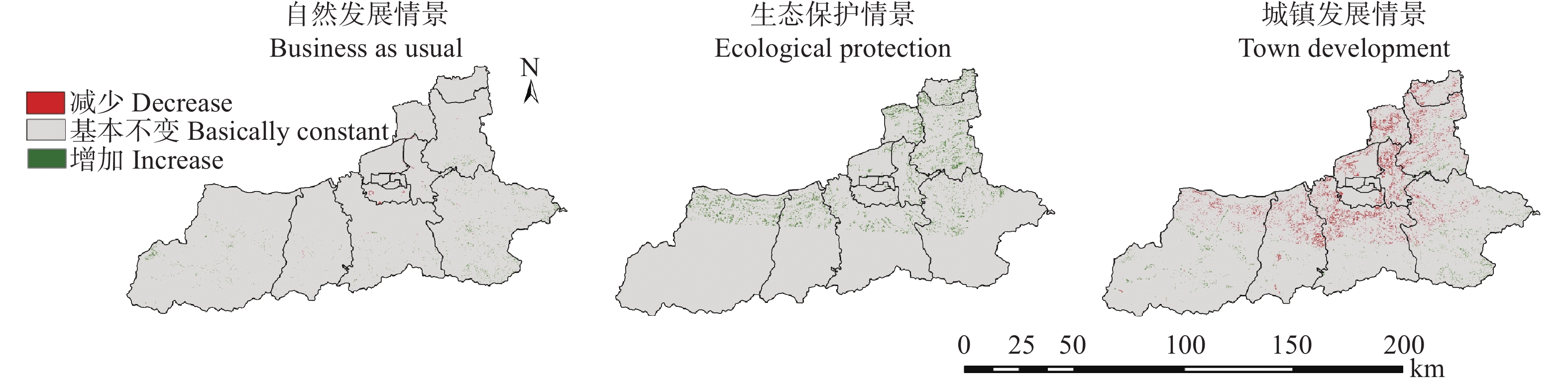
土地利用/覆被变化(LUCC)是陆地生态系统碳储量变化的重要原因, LUCC往往受政策的限制, 从而影响碳储量变化。预测政策指引下的西安市2030年LUCC, 分析其对碳储量的影响, 对西安市政策制定、土地利用结构调整、实现“双碳”目标具有重要意义。本研究基于2000年、2010年和2020年土地利用数据(LULC), 选取11个驱动因子, 根据西安市“十四五”政策规划建立自然发展(Q1)、生态保护(Q2)和城镇发展(Q3) 3个情景, 采用PLUS模型预测并分析西安市2030年土地利用空间分布格局, 并耦合InVEST模型评估西安市在不同发展情景的碳储量变化。研究表明: 1) PLUS模型在西安市的适用性较强, 模型总体精度为0.93, Kappa系数为0.89。2) 2000—2020年西安市建设面积、草地、水体数量增加, 耕地、林地、湿地面积减少, 从转移方向上看, 主要由耕地转为建设用地。3) 2030年, Q1情景延续了以往发展模式, Q2情景下林地、水体等生态用地数量均较2020年有所增加, Q3情景下建设用地大幅增加, 增幅为10.42%。4) LUCC是导致生态系统碳储量变化的主要原因, 2030年Q1情景下碳储量总量较2020年减少373.28 t, 说明延续以往的发展模式会使碳储量总量减少; Q2情景下碳储量总量较2020年增加564.73 t, 说明一定的生态保护措施保护了林地、湿地等生态用地和耕地的数量, 限制了碳密度较高的生态用地和耕地等转化成碳密度较低的建设用地, 可以减缓陆地生态系统碳储量减少趋势, 增加西安市碳储量; Q3情景下碳储量减少734.15 t, 城市化进程的加快, 建设用地规模扩大, 大量的建设用地占用生态用地和耕地, 从而使碳储量大幅减少。研究表明建设用地大幅扩张侵占生态用地和耕地是造成生态系统碳储量流失的主要原因, 实施科学、合理的生态保护措施, 可以很好地解决因经济发展而造成的碳储量下降问题。
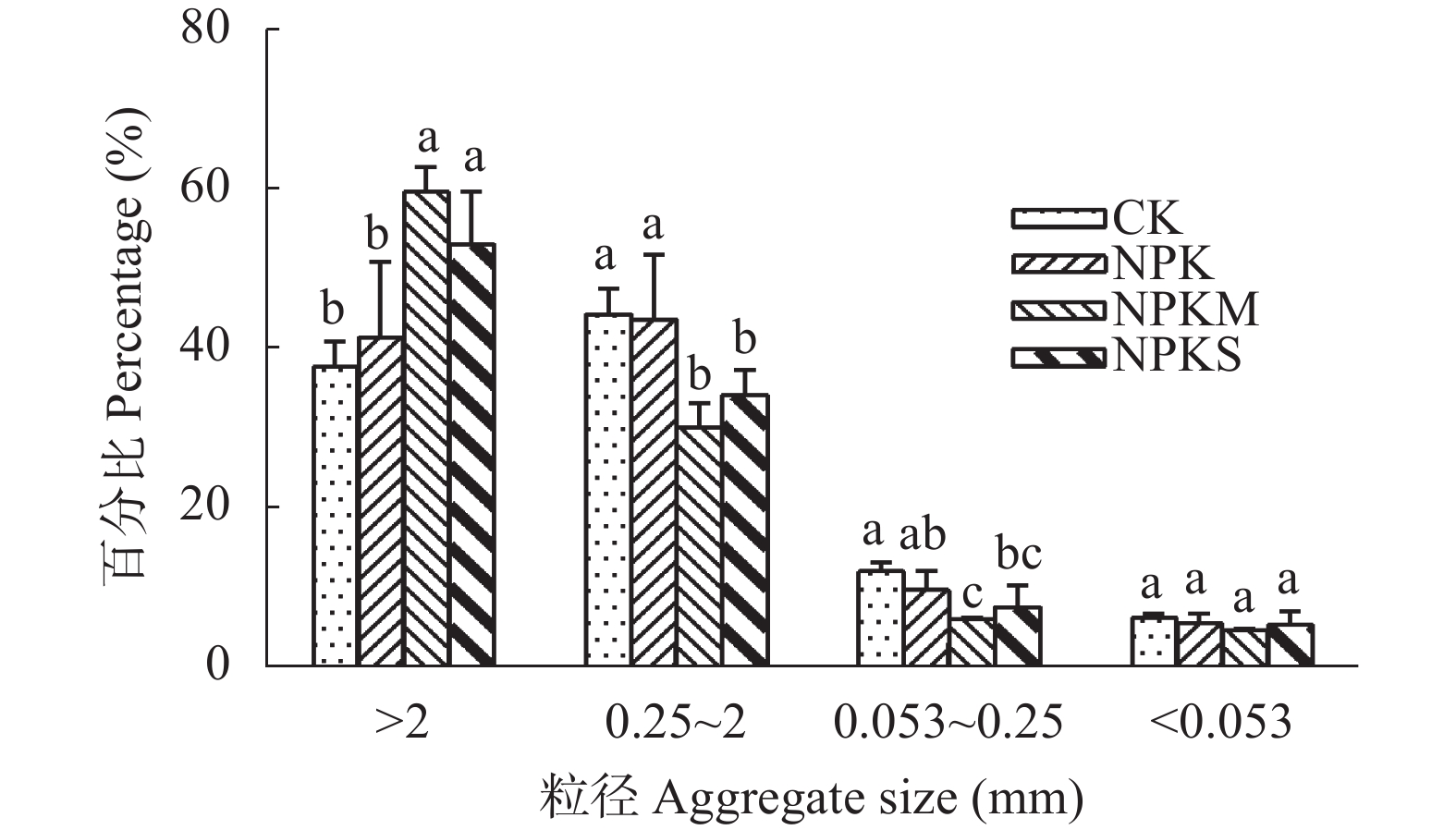
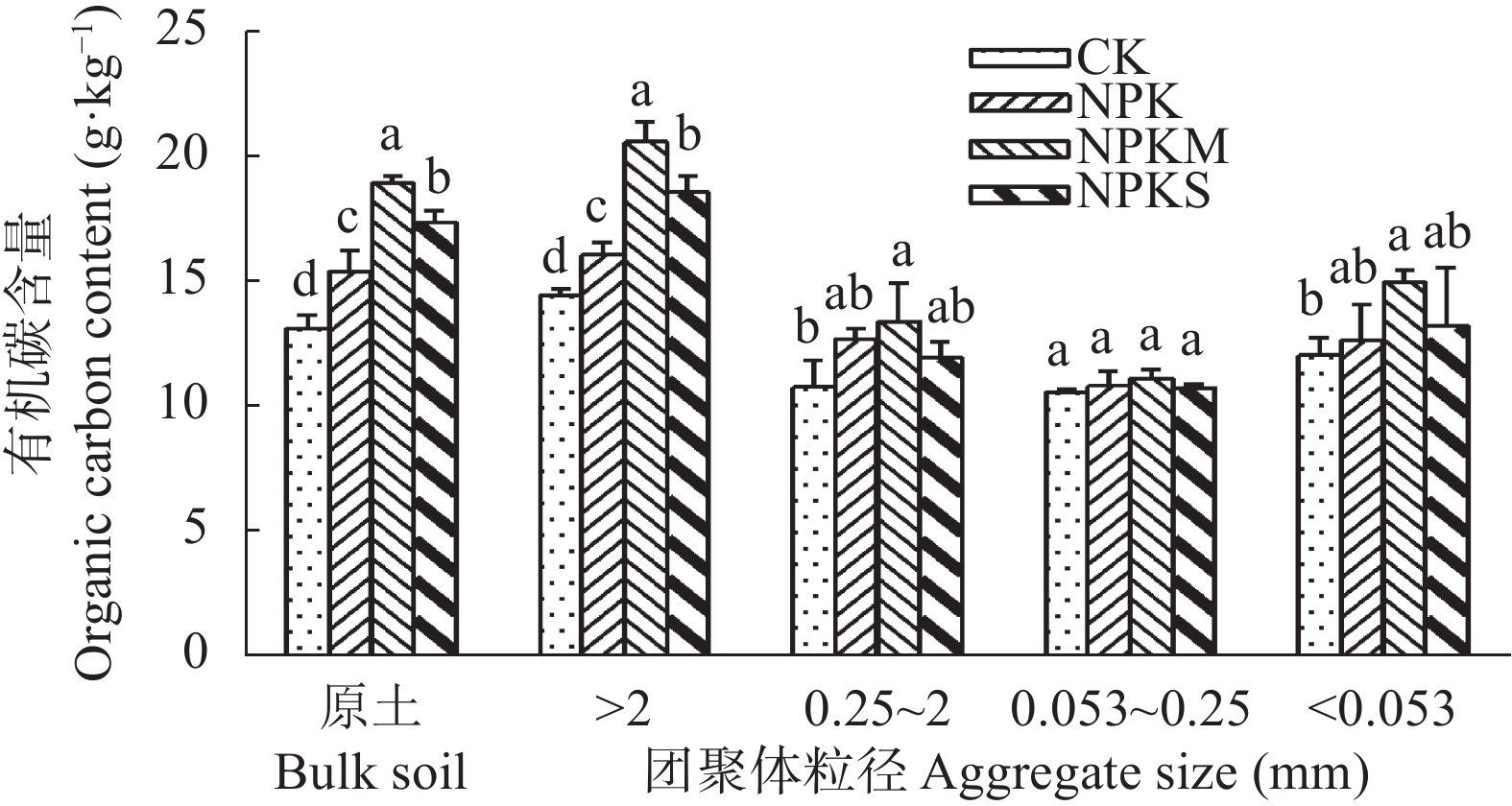
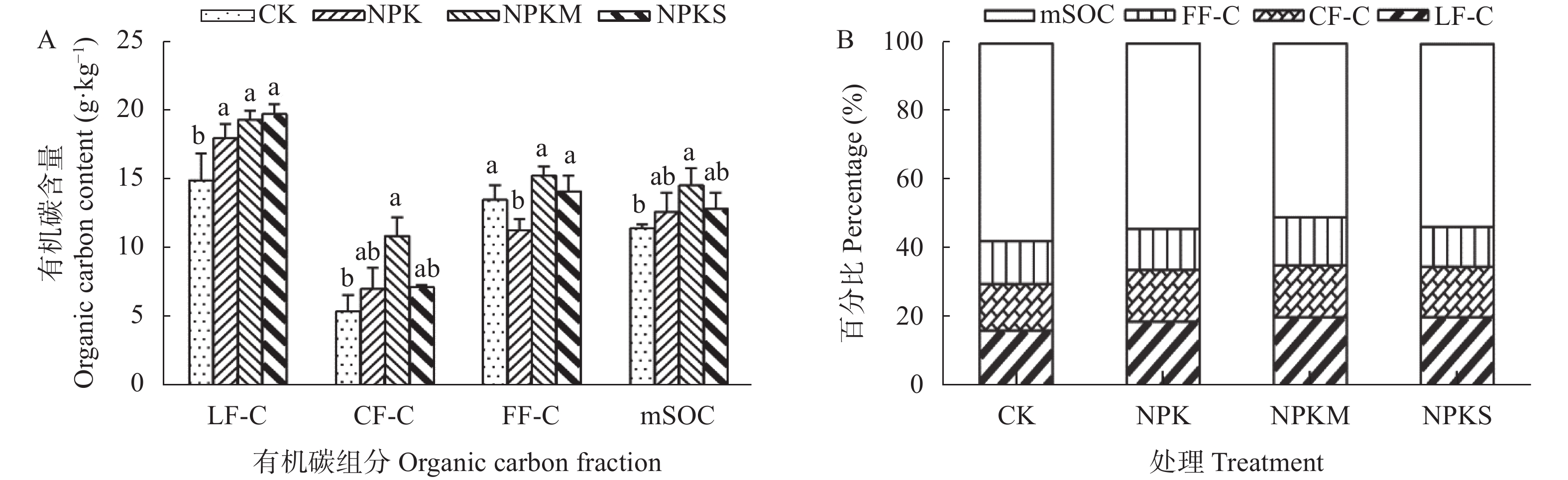
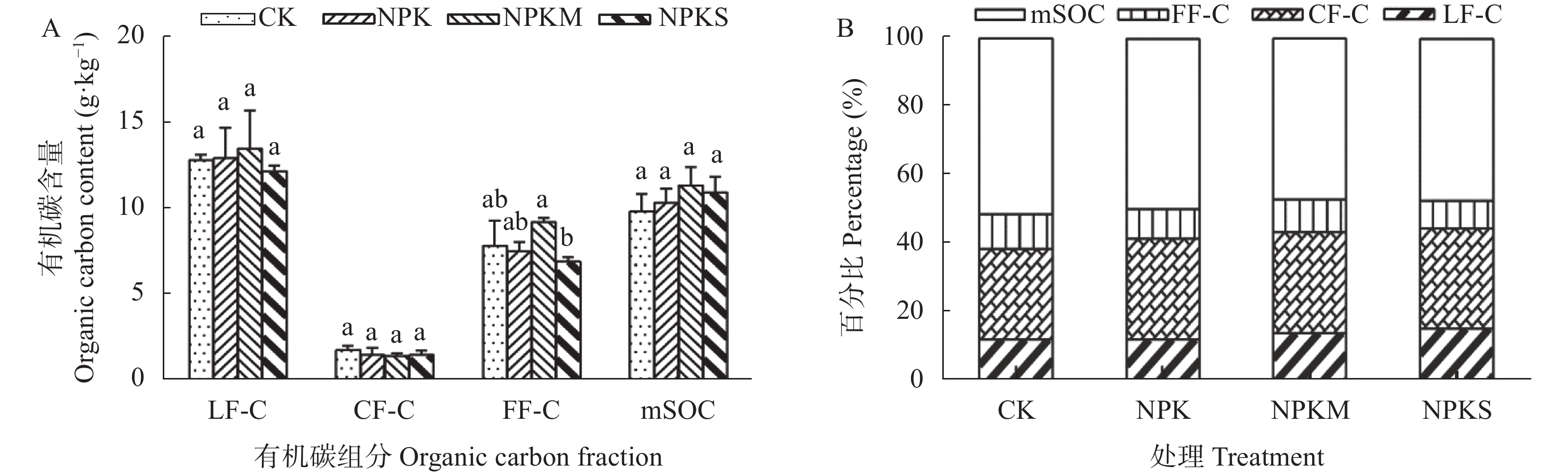
本文旨在研究长期不同施肥处理对南方黄泥田团聚体有机碳固持及其组分分配的影响, 为合理培肥及土壤碳库管理提供依据。基于始于1983年的在黄泥田进行的长期定位试验, 选择不施肥(CK)、单施化肥(NPK)、化肥+牛粪(NPKM)与化肥+全量稻秸还田(NPKS) 4个处理, 采集第36年各处理耕层土壤样品并分析各粒级团聚体有机碳固持及其组分变化。结果表明, 黄泥田耕层土壤以大团聚体(>2 mm)和中间团聚体(0.25~2 mm)为主, NPKM与NPKS处理的土壤大团聚体质量比重分别比CK显著增加22.0和15.5个百分点(P<0.05)。与CK相比, NPKM与NPKS处理中大团聚体对有机碳固持贡献率分别提高25.0和19.3个百分点(P<0.05)。施肥处理的大团聚体内轻组有机碳(LF-C)含量较CK显著增加, 其中NPKS处理后大团聚体中LF-C含量较CK增加32.3% (P<0.05)。大团聚体有机碳含量以及该团聚体内的LF-C含量与水稻产量和有机碳投入量都呈极显著正相关(P<0.01)。以上结果表明, 配施牛粪或秸秆还田有利于增加黄泥田大团聚体比例及其有机碳含量, 进而提高有机碳固持贡献率, 尤其是配施牛粪, 而且有机无机肥配施有利于提高大团聚体内轻组有机碳含量与固持贡献, 秸秆还田更为明显, 可为南方黄泥田施肥管理提供依据。
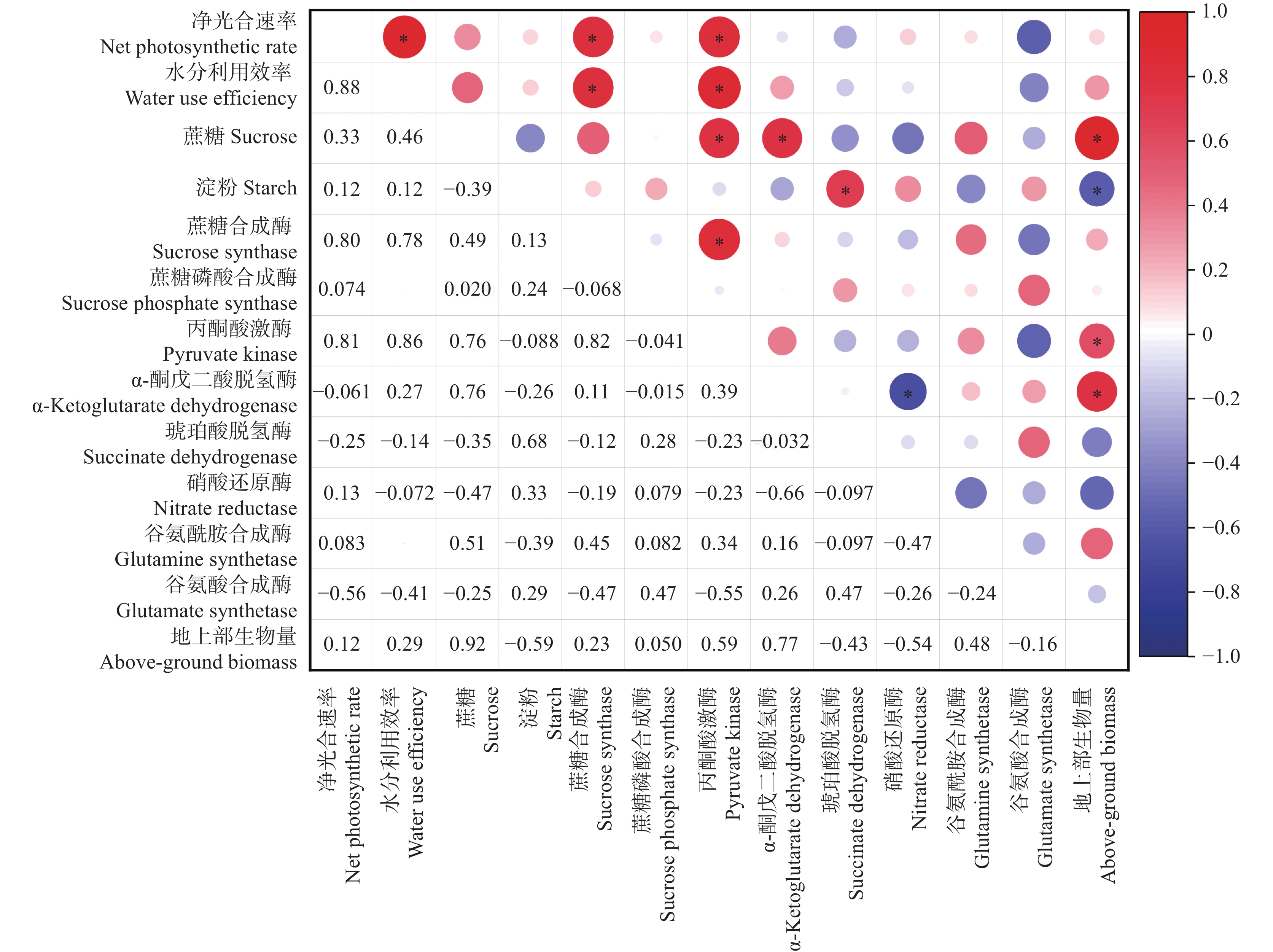
为探讨C4作物玉米对CO2浓度升高、温度升高及其交互作用的响应, 本研究以玉米品种‘先玉335’为材料, 利用人工控制气室设置CK (CO2浓度为400 μmol∙mol−1, 环境温度)、EC (CO2浓度为600 μmol∙mol−1, 环境温度)、ET (CO2浓度为400 μmol∙mol−1, 气温为环境温度+2 ℃)、ECT (CO2浓度为600 μmol∙mol−1, 气温为环境温度+2 ℃) 4个处理, 测定玉米灌浆期叶片光合生理、糖代谢、氮代谢相关指标, 并在成熟后测定玉米生物量。结果表明: 1) CO2浓度升高条件下, 玉米叶片叶绿素含量、蔗糖含量、净光合速率及蔗糖合成酶、丙酮酸激酶和α-酮戊二酸脱氢酶活性显著升高(P<0.05), 但谷氨酸合成酶活性显著降低(P<0.05), 地上部生物量和穗重显著升高35.8%和170.2% (P<0.05)。2)气温升高条件下, 叶片净光合速率、蔗糖合成酶和丙酮酸激酶活性显著升高(P<0.05), 但α-酮戊二酸脱氢酶和谷氨酸合成酶活性显著降低(P<0.05), 地上部生物量、叶重、茎重和穗重显著降低37.0%、28.7%、32.3%和62.2% (P<0.05)。3) CO2浓度和气温均升高条件下, 叶片净光合速率和丙酮酸激酶活性显著升高(P<0.05), 但叶绿素含量、α-酮戊二酸脱氢酶和谷氨酸合成酶活性显著降低(P<0.05), 叶重显著降低23.4% (P<0.05)。总之, CO2浓度升高可通过促进玉米叶片光合速率, 增加糖代谢相关酶活性和光合代谢产物等缓解温度升高对玉米生物量的负效应; CO2浓度升高、气温升高以及二者互作下玉米氮代谢受到抑制, 玉米叶片受到氮素胁迫, 或对玉米品质产生不利影响。
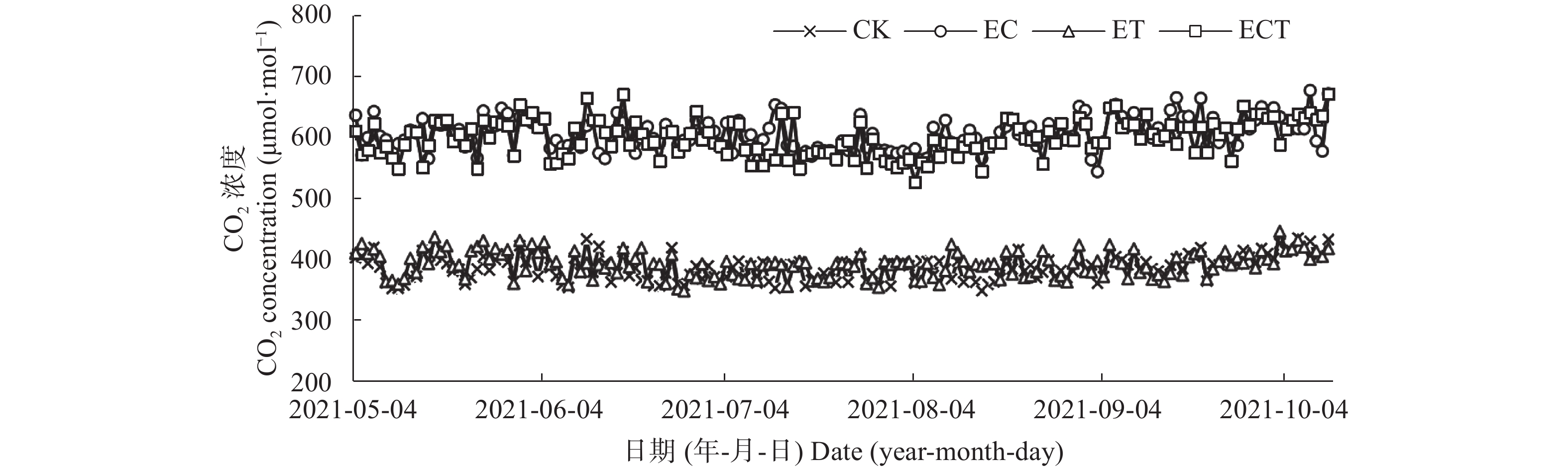
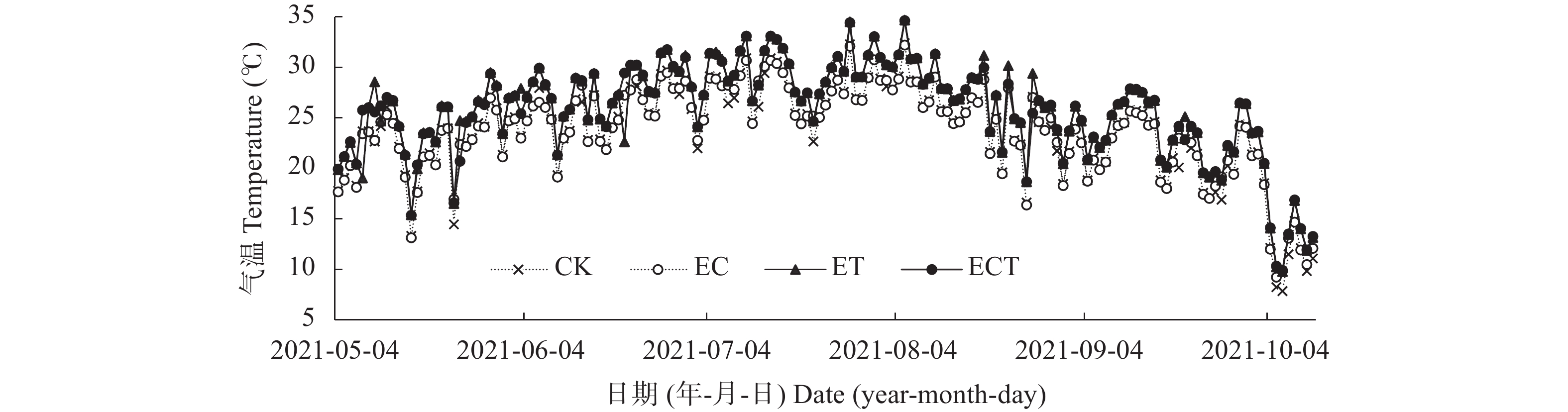
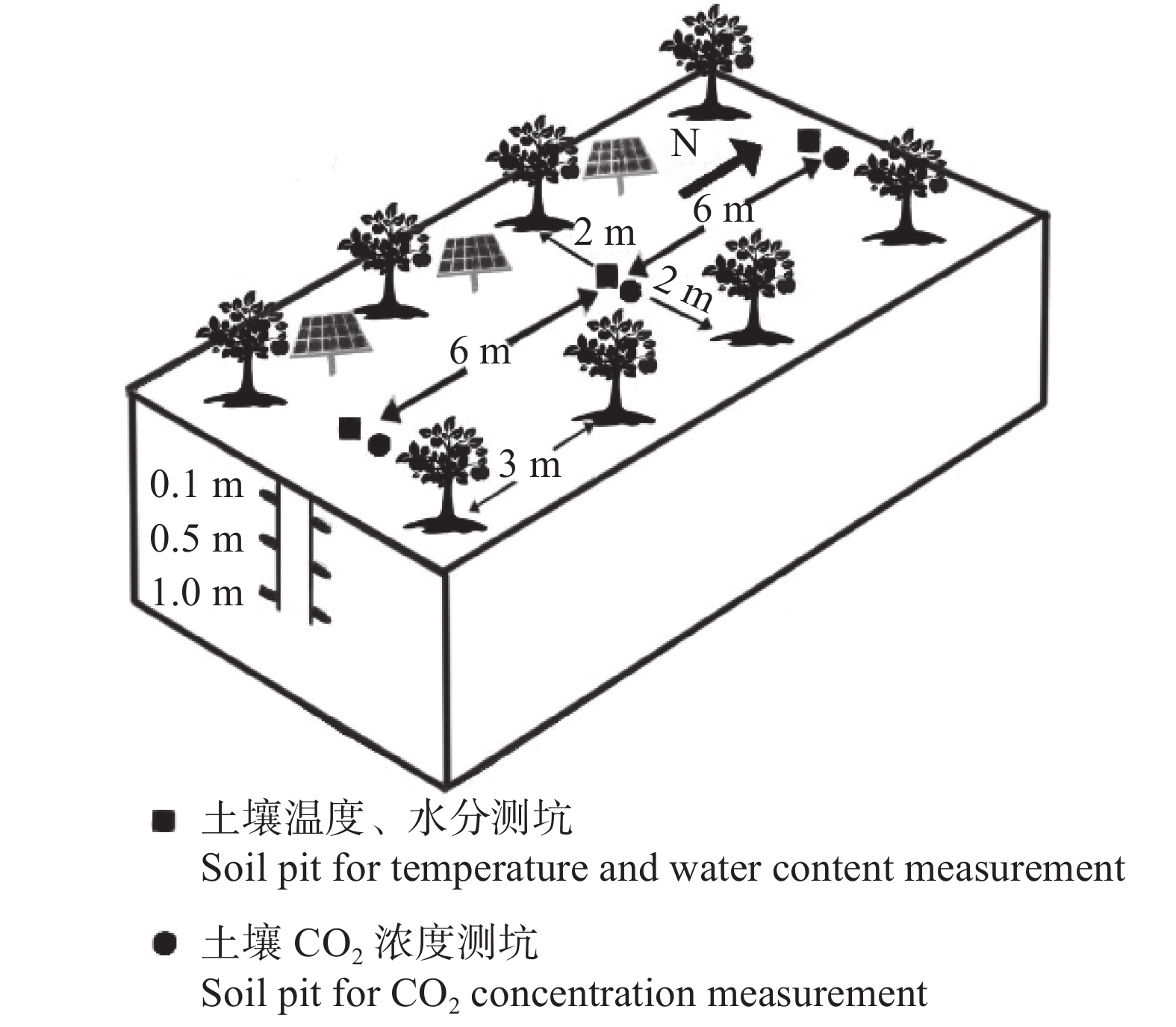
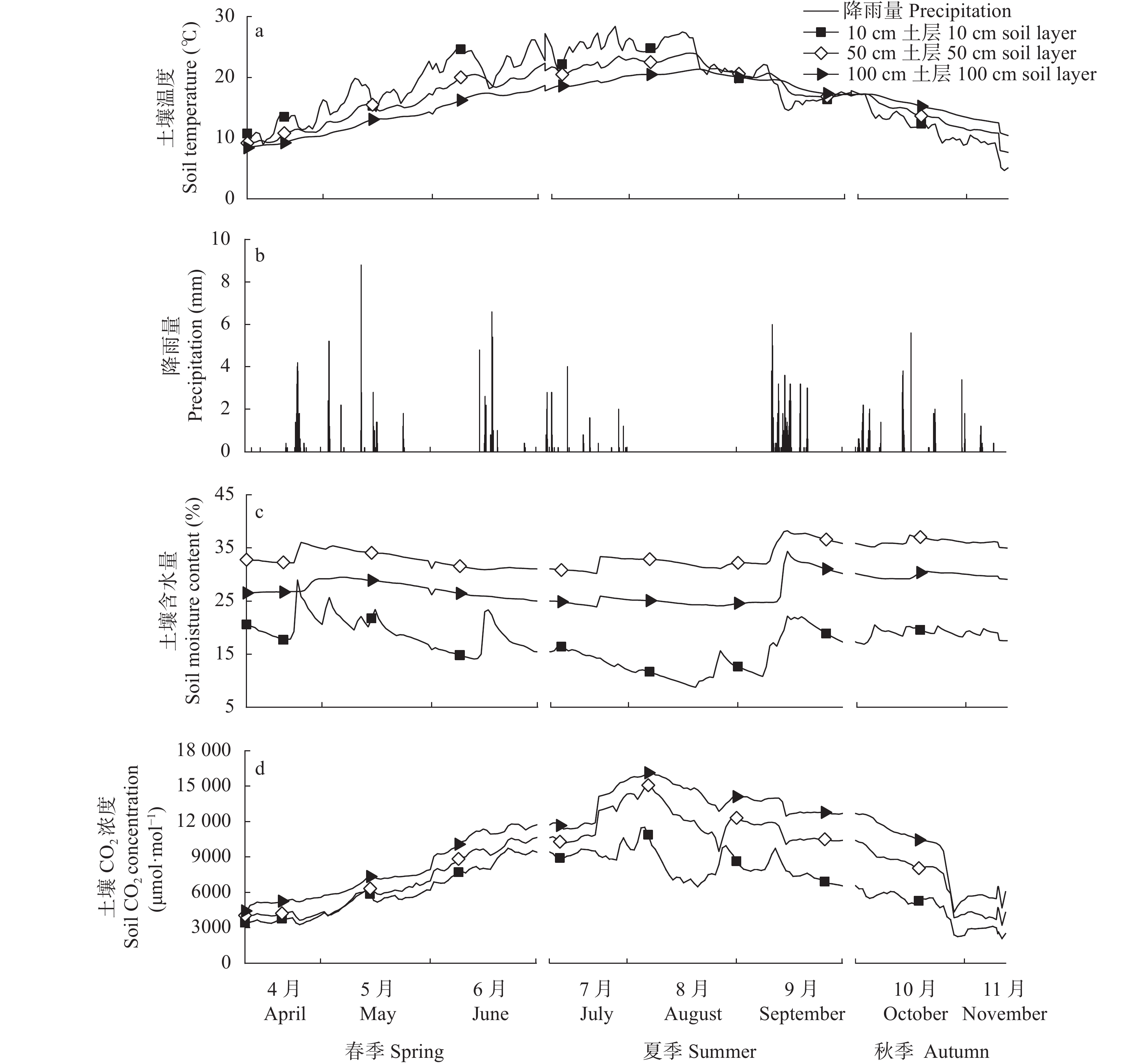
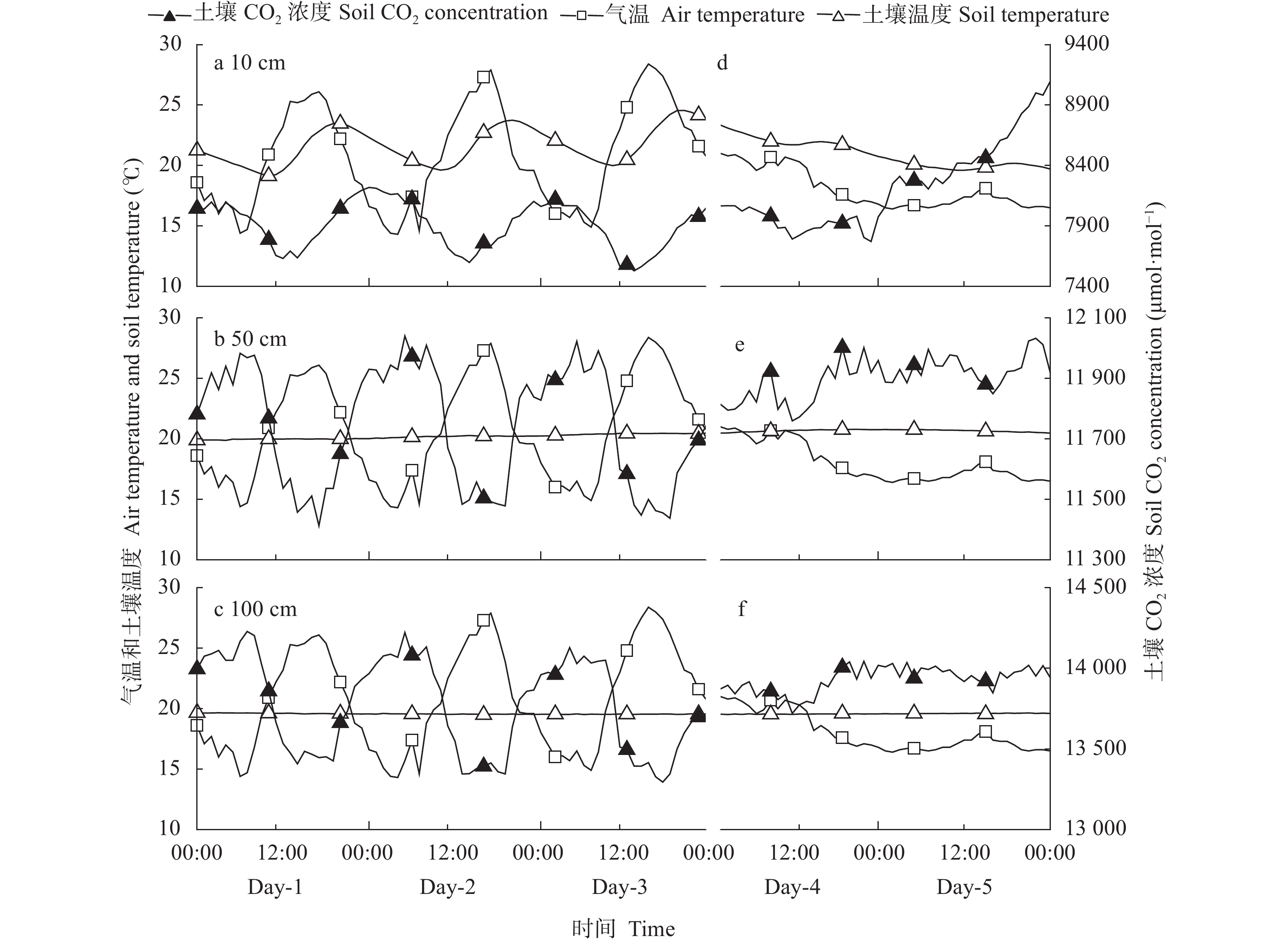
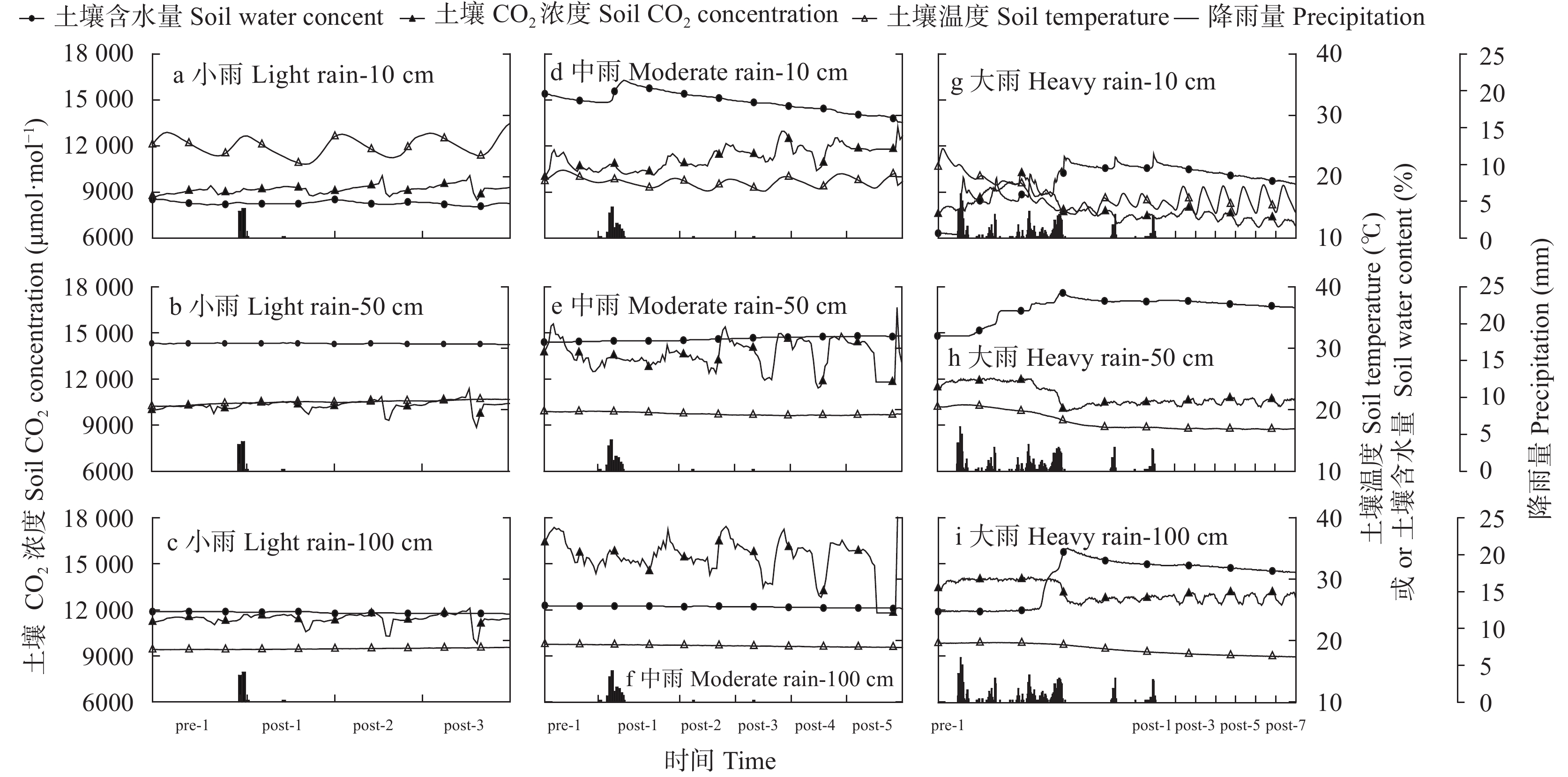
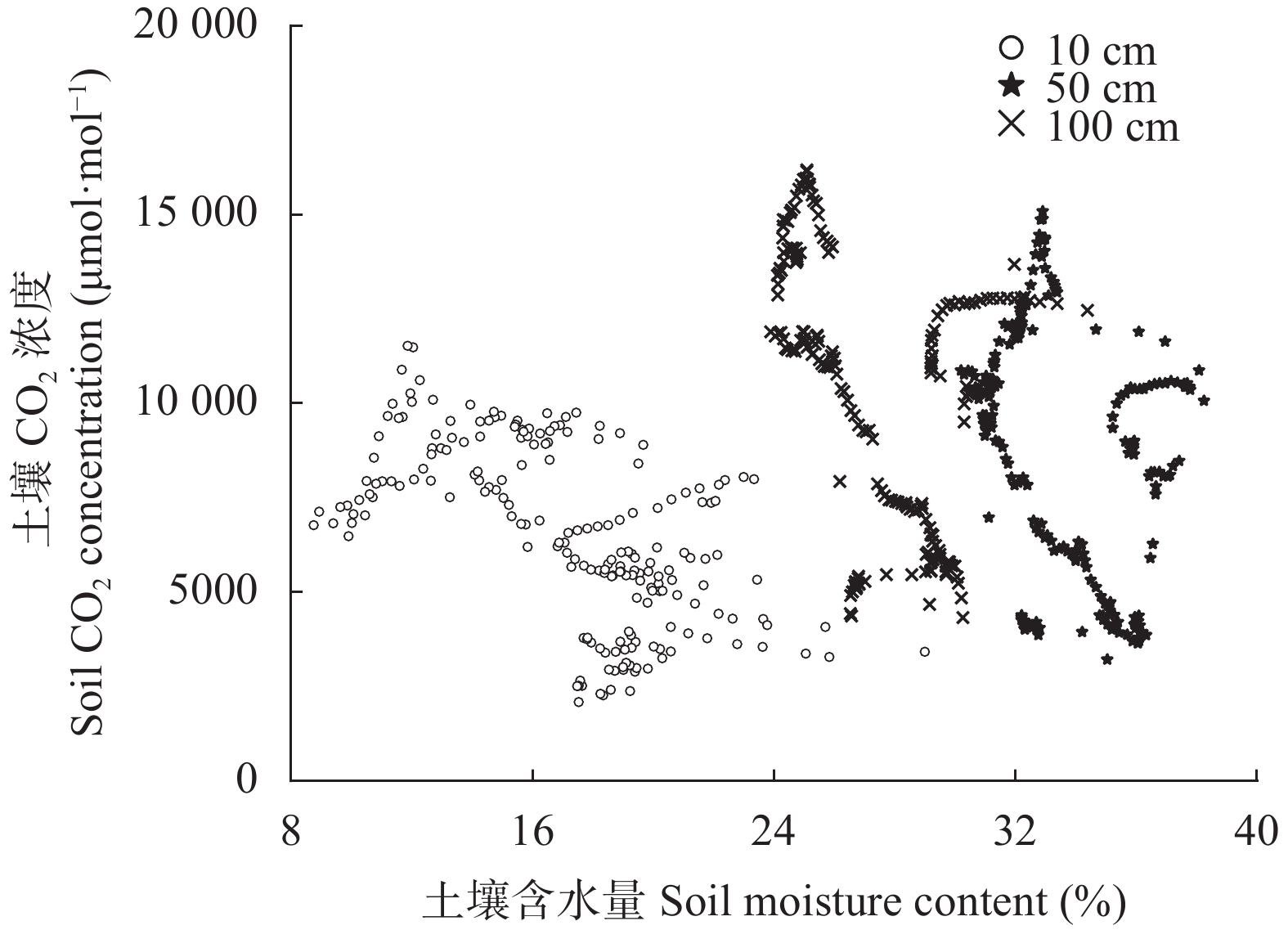
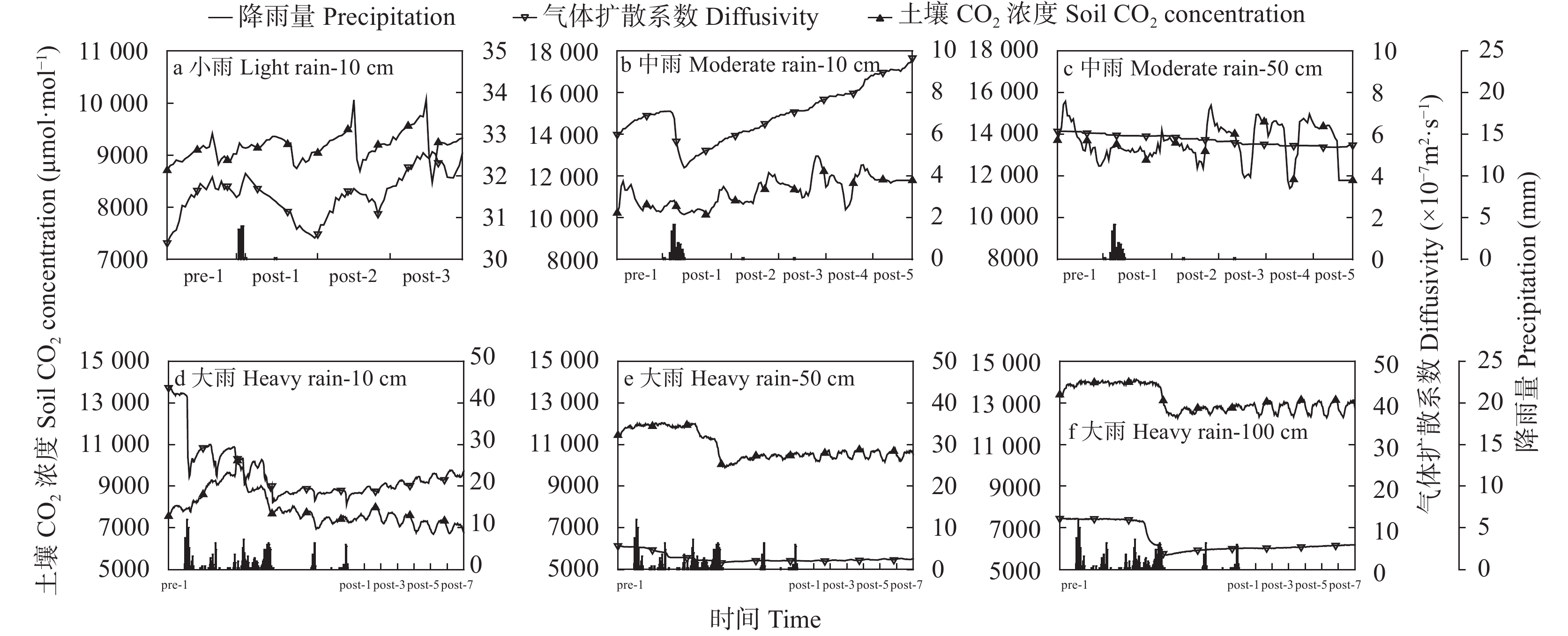
降雨是干旱半干旱地区土壤CO2产生、传输或扩散的重要影响因素, 并进一步影响土壤和大气中的CO2浓度。目前大量研究集中在地表CO2通量变化与降雨的关系, 深层土壤有机碳储量巨大, 但深层土壤CO2浓度变化对降雨事件的响应机制尚不清楚。本研究通过对10 cm、50 cm和100 cm处土壤CO2浓度进行原位连续监测, 分析不同深度土壤CO2浓度对降雨事件的响应过程及其影响因素。结果表明: 试验期间, 78%的降雨事件能迅速引起10 cm处土壤CO2浓度发生改变, 且随着降雨量增大, 土壤CO2浓度发生变化的深度逐渐增加。当降雨量在10~25 mm时, 50 cm处土壤CO2浓度在91 h后降低; 降雨量>25 mm时, 100 cm处土壤CO2浓度在121 h后降低。当土壤由干变湿时, 降雨量>25 mm的降雨事件促进10 cm处土壤CO2浓度升高30%后开始降低, 而50 cm和100 cm处土壤CO2浓度随水分升高分别降低16.3%和10.9%。在半干旱区, 当土壤含水量较低时, 降雨可以对10 cm处土壤CO2浓度变化产生短暂的正激发效应, 而深层土壤含水量往往高于田间持水量, 水分升高会导致该处土壤CO2浓度降低。降雨对不同深度土壤CO2浓度变化的影响存在差异, 这在很大程度上取决于土壤含水量状况。